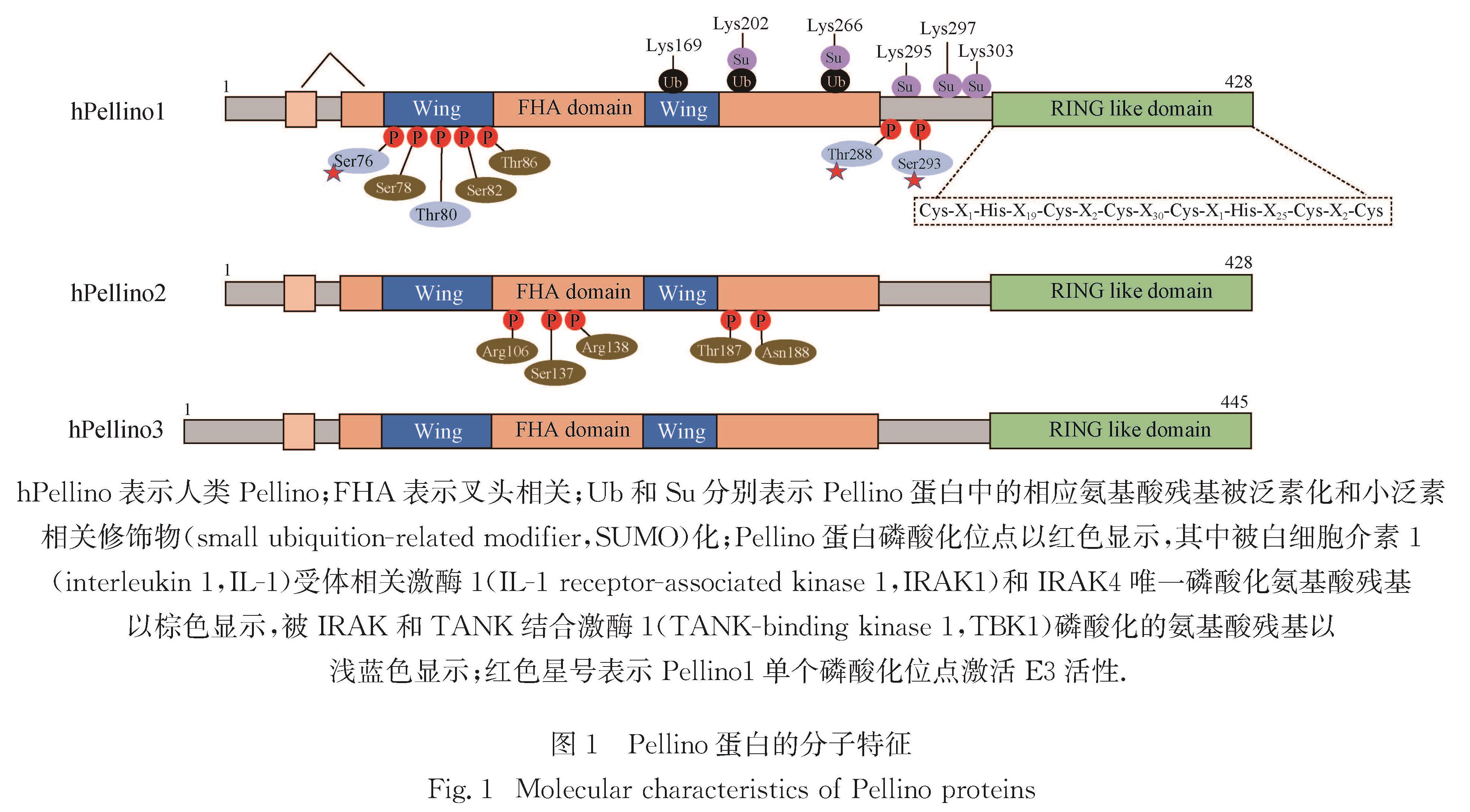
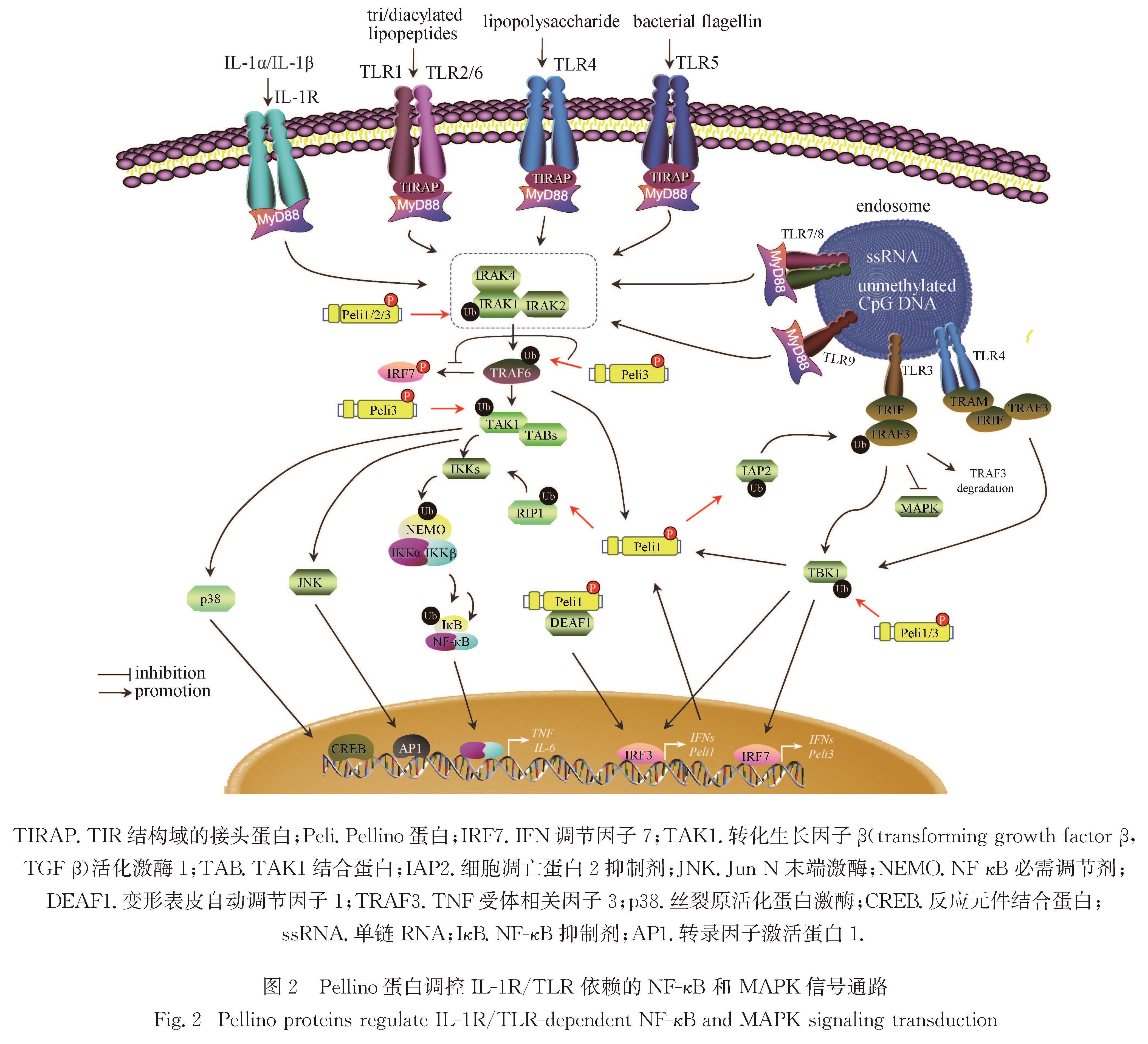
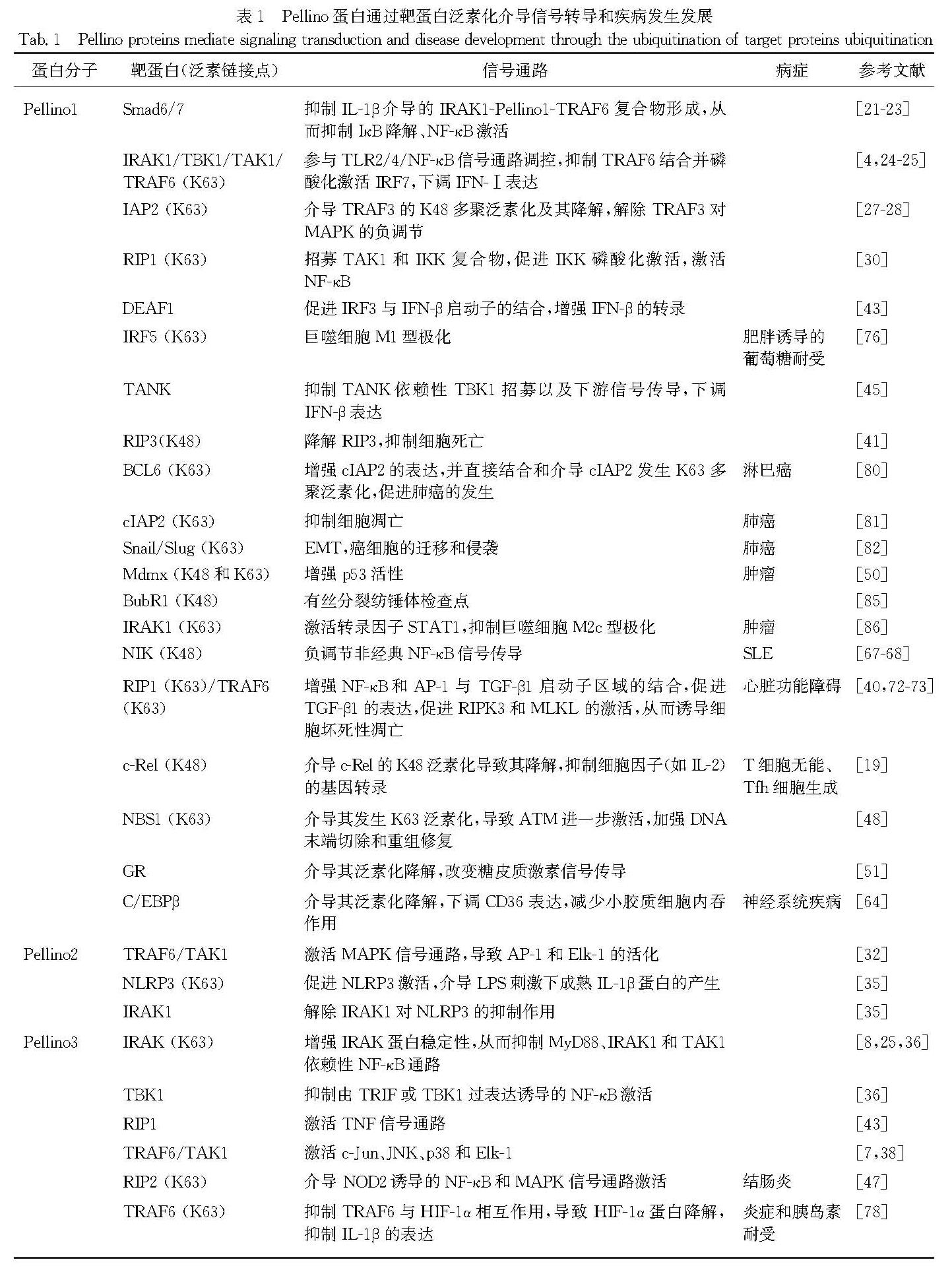
2.1.1 Pellino蛋白调控IL-1R/TLR依赖的NF-κB和MAPK信号转导
固有免疫系统利用独特的模式识别受体(pathogen recognition receptors,PRRs)识别外源性病原体相关分子(pathogen-associated molecular patterns,PAMPs).TLRs是PRRs的关键成员,其识别PAMPs后招募含TIR(Toll/IL-R)结构域的接头蛋白,包括髓样分化因子88(myeloid differentiation primary response protein 88,MyD88)和干扰素β(interferon β,IFN-β)TIR结构域衔接蛋白(TIR domain-containing adapter protein inducing IFN-β,TRIF).MyD88是TLRs所有成员(除TLR3外)共有的接头蛋白,而TRIF主要参与TLR3和TLR4介导的信号通路; TLR3直接招募TRIF,而TLR4以TRIF相关接头分子(TRIF-related adapter molecule,TARM)作为支架蛋白间接招募TRIF.IL-1受体(IL-1 receptor,IL-1R)/TLR信号途径通过激活下游NF-κB、MAPK等信号共同诱导促炎细胞因子和趋化因子的表达(图2).
IL-1结合IL-1R后刺激Pellino1与IRAK4、IRAK1和TRAF6之间的相互作用,激活MyD88依赖的NF-κB信号和IL-8基因表达[5].TGF-β刺激下,Pellino1与Smad6或者Smad7相互作用,竞争性阻止IL-1β介导的IRAK1-Pellino1-TRAF6复合物形成,从而抑制IκB降解、NF-κB激活以及促炎因子表达[21-23].在LPS刺激下,Pellino1介导IRAK1、TBK1、TAK1和TRAF6的K63多聚泛素化,参与TLR2/4/NF-κB信号通路调控[24-25].敲低Pellino1可抑制炎性细胞因子、趋化因子和黏附分子的表达以及NF-κB RelA的转录活性[26].此外,小胶质细胞中Pellino1介导IAP2的K63多聚泛素化可增强其蛋白稳定性,激活的IAP2介导TRAF3的K48多聚泛素化导致其降解,从而解除TRAF3对MAPK的负调节[27-28].然而,Pellino1介导的NF-κB和MAPK信号转导仅限于免疫细胞,在非免疫性细胞中未发现相似的调控作用[29-32].
图2 Pellino蛋白调控IL-1R/TLR 依赖的NF-κB 和 MAPK信号通路
Fig.2 Pellino proteins regulate IL-1R/TLR-dependent NF-κB and MAPK signaling transduction
有研究表明,Pellino1通过介导受体相互作用蛋白1(receptor-interacting protein 1,RIP1)的K63多聚泛素化,招募TAK1和IKK复合物,促进IKK复合物的磷酸化激活,最终导致NF-κB的激活和促炎细胞因子的增加; 敲除Pellino1能显著抑制TLR3/4配体对炎性细胞因子的诱导,并使小鼠对感染性休克具有更强的抵抗力[30].而另一项研究发现Pellino1缺失对聚肌胞苷酸(polyinosinic-polycytidylic acid,poly(I:C))诱导的RIP1泛素化、IKK和MAPK信号通路激活以及炎性细胞因子表达均没有影响[33].因此,Pellino1是否介导TRIF依赖的NF-κB和MAPK信号转导有待进一步明确.
在LPS刺激的RAW264.7细胞中,B细胞淋巴瘤10(B cell leukemia/lymphoma 10,BCL10)与TLR4结合,随后与Pellino2相互作用以激活NF-κB通路,但是Pellino2缺失并不能完全消除BCL10对NF-κB的激活作用,表明Pellino2部分参与BCL10激活NF-κB信号通路[34].另一项研究也发现,敲低Pellino2不能完全抑制IL-1和LPS诱导的炎症反应[11].此外,Pellino2过表达不能激活NF-κB信号转导,但可增强其下游基因的表达[6,25].这些研究表明,Pellino2在IL-1R/TLR依赖的NF-κB信号转导中的调控作用有待深入探讨.Pellino2是否调控IL-1R/TLR依赖的MAPK信号通路也存在争议.Pellino2与TRAF6和TAK1相互作用,介导其泛素化,从而激活MAPK信号通路,导致AP-1和Elk-1(Ets-like protein1)的活化[32].在Pellino2敲低的293细胞和巨噬细胞中,IL-1和LPS介导的JNK和胞外信号调节激酶(extracellular regulated protein kinase,ERK)磷酸化减弱[11].然而,在其他研究中发现,Pellino2缺失并不影响IL-1和LPS介导的MAPK信号通路的激活[16,35].由于序列和结构具有高度同源性,Pellino蛋白成员之间可能存在功能性代偿,导致Pellino2缺失细胞中MAPK信号通路变化不明显.
Pellino3b通过介导IRAK的K63多聚泛素化竞争性抑制K48多聚泛素化,增强IRAK蛋白稳定性,从而抑制MyD88、IRAK1和TAK1依赖性NF-κB通路[8,25,36].Pellino3b也可通过介导TBK1的泛素化,抑制由TRIF或TBK1过表达诱导的NF-κB激活[36].敲除Pellino3导致LPS依赖性炎性细胞因子IL-1β 的表达上调[20].这些研究表明Pellino3负向调节IL-1R/TLR依赖的NF-κB信号通路.此外,也有研究表明Pellino3正向调节IL-1R/TLR依赖的MAPK信号通路,Pellino3通过介导TRAF6和TAK1的泛素化,诱导c-Jun、JNK、p38和Elk-1的激活[7,37].在Pellino3敲减细胞中,IL-1诱导的MAPK信号通路激活被减弱[38].
2.1.2 Pellino蛋白调控TNF信号通路
TNF既可以激活NF-κB以诱导炎性和抗凋亡蛋白的表达,也可以触发caspase级联反应促进细胞凋亡.TNF与其受体TNFR1结合后,TNFR1发生多聚化并与细胞质中TNFR1相关死亡结构域蛋白(TNFR1-associated death domain protein,TRADD)相互作用,招募RIP1、TRAF2或TRAF5以及细胞凋亡抑制蛋白,形成复合物Ⅰ.TRAF2和IAP可导致RIP1的K63多聚泛素化以增强其稳定性,然后招募 IKK 复合物和TAK1,最终激活NF-κB和抗凋亡基因的表达.同时,NF-κB也会上调A20和肿瘤抑制因子CYLD的表达,对RIP1去泛素化以解除其激活NF-κB 的作用.此外,未泛素化的RIP1可以与TNFR1 分离,与Fas相关死亡域(Fas-associated death domain protein,FADD)蛋白相互作用,该蛋白反过来招募procaspase-8以形成细胞内信号传导复合物Ⅱ.复合物Ⅱ进而触发caspase-8的酶切加工和激活,启动caspase级联反应.更多细节和相关进展可参阅文献[39].
Pellino1是TNF-α介导的细胞死亡途径的关键调节剂,可以促进坏死性凋亡而抑制细胞凋亡.一方面,Pellino1介导受体相互作用蛋白激酶1(receptor-interacting protein kinase 1,RIPK1)的K63多聚泛素化,增强其与RIPK3的结合以促进RIPK3和MLKL(mixed lineage kinase domain-like protein)的激活,从而诱导细胞坏死性凋亡; 另一方面,Pellino1的缺失通过上调c-Myc的表达和下调caspase-8抑制分子c-FLIP(cellular FLICE inhibitory protein)的表达,使细胞对TNF-α诱导的RIPK1依赖性和RIPK1非依赖性细胞凋亡更敏感[40].敲减Pellino1也导致TNF刺激下NF-κB RelA(p65)的转录活性及其下游促炎细胞因子、趋化因子和黏附分子的表达下调[26].另一项研究发现,TNF-α刺激的293T或宫颈癌HeLa细胞中,Pellino1可介导RIP3的K48多聚泛素化,导致其降解并抑制细胞死亡[41].这些研究说明Pellino1在不同的细胞背景中参与调控TNF信号转导的分子机制存在差异.
Pellino3也可以介导RIP1的泛素化,进而激活TNF诱导的复合物Ⅰ介导的NF-κB信号通路,发挥促炎抗凋亡作用以促进细胞存活; 同时抑制TNF诱导的复合物Ⅱ形成,抑制caspase-8介导的细胞凋亡[42].
2.1.3 Pellino2调控炎性小体信号通路
炎性小体是一种由NOD样受体热蛋白结构域相关蛋白(NOD-like receptor thermal protein domain associated protein,NLRP)、凋亡相关斑点样蛋白(apoptosis-associated speck-like protein containing a CARD,ASC)和半胱天冬酶-1前体(pro-cysteinyl aspartate specific proteinase-1,pro-caspase-1)组成的胞浆多蛋白复合物,能被多种病原相关分子模式或损伤相关分子模式激活.NLRP3炎性小体将接头蛋白ASC和caspase-1募集到寡聚复合物中,导致pro-caspase-1自动蛋白水解加工成活性形式,将pro-IL-1β和pro-IL-18切割为成熟的分泌形式,引发炎性反应和细胞焦亡.研究证实 Pellino2的FHA和RING结构域通过促进NLRP3的K63泛素化来促进NLRP3激活,介导LPS刺激下成熟IL-1β蛋白的产生; Pellino2缺陷小鼠的骨髓细胞在TLR启动、NLRP3 刺激和细菌攻击时,NLRP3的激活受损,对LPS刺激也有很强的耐受性; 此外,Pellino2 还可通过泛素化IRAK1解除IRAK1对NLRP3的抑制作用[35].
2.2 Pellino蛋白参与免疫反应调节
2.2.1 Pellino蛋白调控IFN-Ⅰ的表达
IRF是调控IFN基因表达的关键转录因子,在抗感染免疫中发挥重要作用.小鼠胚胎成纤维细胞中Pellino1缺失不影响TLR3/4诱导的IKKε激活和IFN-β表达,提示Pellino1不参与调控TRIF依赖的IRF3激活和随后的IFN-β表达[30].然而,在病毒双链RNA的刺激下,Pellino1敲除小鼠的骨髓细胞和成纤维细胞中IFN-β表达下调,IRF3与IFN-β基因启动子的相互作用受损,表明Pellino1对病毒感染等因素刺激下的IFN-β基因表达具有重要调控作用[33].进一步研究证实Pellino1与转录因子DEAF1同源物存在相互作用,两者促进IRF3与IFN-β启动子的结合,促进IFN-β的转录[43].研究发现Pellino2 缺陷的树突状细胞经TLR9 激活后,小鼠IFN-Ⅰ的产生受损[44].
Pellino3通过两种途径参与IFN-Ⅰ的表达调控:1)介导TRAF6的泛素化,抑制其结合并磷酸化激活IRF7,导致IFN-Ⅰ表达下调[4]; 2)巨噬细胞中LPS激活TLR4后,TRIF依赖的信号级联将TRAF3、TANK、TBK1组装成复合体,进而磷酸化IRF3,磷酸化的IRF3易位到细胞核中,与其反应元件结合并激活IFN-β和IP-10基因的转录.当巨噬细胞中加入氧化型低密度脂蛋白后,上调的Pellino3通过介导TANK的单泛素化,抑制TANK依赖性TBK1招募以及下游信号传导,从而抑制IFN-β表达[45].
2.2.2 Pellino1调控T细胞活化
T细胞活化需要两个信号刺激,即TCR和共刺激信号分子(如CD28).T细胞中TCR和CD28的共刺激信号招募IKK复合物,催化IκB蛋白的磷酸化,导致其降解以及NF-κB家族Rel亚基的释放,游离c-Rel 易位至细胞核中增强CD28效应分子的转录(如IL-2),从而驱动T细胞的活化和增殖[38].
Pellino1在B细胞和T细胞中高表达,其敲除导致T细胞过度活化,并使T 细胞难以被调节性T细胞和TGF-β抑制[19].Pellino1作为T细胞活化的关键负调节因子可缓解自身免疫,Pellino1敲除小鼠可自发地发生以多器官炎症和自身抗体产生为特征的自身免疫疾病[19].TCR和CD28的共刺激信号增强Pellino1表达,后者通过介导c-Rel K48泛素化导致其降解,抑制细胞因子(如IL-2)表达,最终导致一种与外周T细胞耐受相关的效应称“T细胞无能”[19].此外,有研究发现Pellino1通过介导c-Rel的泛素化调控滤泡辅助性T(T follicular helper,Tfh)细胞的生成和功能.miR-155通过靶向抑制Pellino1的表达,进而抑制NF-κB家族转录因子c-Rel的降解,抑制Tfh细胞增殖和CD4+ T细胞上的CD40配体表达[46].
2.2.3 Pellino3调控NOD2信号通路
Pellino3是NOD2信号通路的重要调控因子,通过介导激酶RIP2的泛素化参与肠道炎症反应[47]:细菌侵入肠上皮细胞产生肽聚糖衍生肽(如胞壁酰二肽),触发NOD2 的寡聚化,活化的NOD2招募蛋白激酶RIP2.Pellino3通过FHA结构域与RIP2相互作用并介导其K63多聚泛素化,促使TAK1和IKK复合物的共同招募,激活IKK和下游NF-κB 信号通路.TAK1还可以激活MAPK信号通路(p38、JNK和ERK),从而激活AP-1等转录因子,与NF-κB协同诱导细胞因子、趋化因子和杀菌肽的表达.
2.3 Pellino蛋白参与其他信号通路调控
DNA双链断裂(DNA double-strand break,DSB)和重组修复对于维持基因组完整性至关重要.DSB触发DNA损伤反应信号,激活DNA损伤修复和细胞周期检测点机制.多项研究证实Pellino1参与DNA损伤修复信号通路的调控:DNA损伤传感蛋白激酶ATM(ataxia-telangiectasia mutated gene)招募并磷酸化Pellino1到DSB位点,磷酸化组蛋白H2AX促使Pellino1与NBS1(Nijmegen breakage syndrome 1)相互作用并介导其发生K63泛素化,导致ATM的进一步激活,从而加强DNA末端切除和重组修复; DNA损伤后也激活p53信号通路及其靶基因p21的转录,形成细胞周期G1检查点,以减少受损DNA的复制和积累[48].另一项研究发现,Pellino1通过FHA结构域与磷酸化的p53相互作用介导其转录活化[49].Pellino1也可以通过与p53抑制蛋白Mdmx相互作用介导其泛素化,从而抑制其细胞核易位,解除其对p53的抑制作用[50].
此外,Pellino1也参与G蛋白偶联受体信号转导的调控.骨架蛋白β-arrestin-1被证实是一个新的糖皮质激素受体(glucocorticoid receptor,GR)互作蛋白,敲低后促进Pellino1表达; Pellino1与GR相互作用并介导其泛素化降解,改变糖皮质激素信号传导[51].