2.1 红树林根际沉积物中分离和筛选的解磷真菌
以磷酸钙为唯一磷源时,从福建、海南、台湾三地的红树林根际沉积物中分离和筛选出13株解磷真菌,并进一步分析这些真菌菌株对PR的解磷能力.由表1可知,与未接种处理相比,菌株PSF-TW表现出最强的解磷能力,高达(160.77±6.92)mg/L,其次为菌株PSF-FJ1,解磷能力为(155.41±2.73)mg/L.伴随着菌株的生长和可溶性磷酸盐含量的变化,所有菌株都使得培养液的pH明显降低.Achal等[24]的研究指出,
表1 解磷真菌在含PR的NBRIP液体培养基中的解磷能力
Tab.1 phosphate-solubilizing capacity of phosphate-solubilizing fungi in NBRIP liquid medium with PR
利用紫外线照射获取的A. tubingensis表型突变体AtM-5和AtM-2,可分泌酸性磷酸酶和植酸酶来降低培养液的pH,从而有效溶解PR.从印度奥里萨邦重金属矿区中筛选出的解磷真菌Penicillium sp.21和Penicillium sp.2,与解磷细菌相比具有更高的产酸效率,也表现出更强的PR溶解能力[25].
2.2 解磷真菌在SLP固体培养基中对Pb的耐受能力
除了解磷能力之外,菌株的重金属耐受能力在重金属污染水体修复中亦至关重要.如表2所示,在SLP固体培养基中,当Pb质量浓度低于200 mg/L时,重金属Pb对菌株的抑制效果不明显,说明供试菌株对Pb具有一定的耐受能力; 但当Pb质量浓度达到1 000 mg/L时,菌株PSF-FJ2、PSF-FJ7和PSF-FJ9已无法生长,菌株PSF-FJ4、PSF-FJ10和PSF-FJ11的生长则受到明显抑制; 菌株PSF-FJ3、PSF-FJ1、PSF-TW和PSF-HN对Pb的耐受能力最强,甚至在Pb质量浓度高达2 500 mg/L时仅受到轻微抑制.Babu等[26]从含Pb尾矿中筛选出耐受能力强的青霉属Penicillium sp.PDR1~3、镰刀菌属Fusarium sp.PDR1~16和木霉属Trichodermasp.PDR1~7,它们在固体培养基中对Pb的MIC范围为1 700~2 100 mg/L.
表2 不同Pb质量浓度对SLP固体培养基中解磷真菌生长情况的影响
Tab.2 Influence of different Pb concentrations on growth of phosphate-solubilizing fungi on SLP solid medium
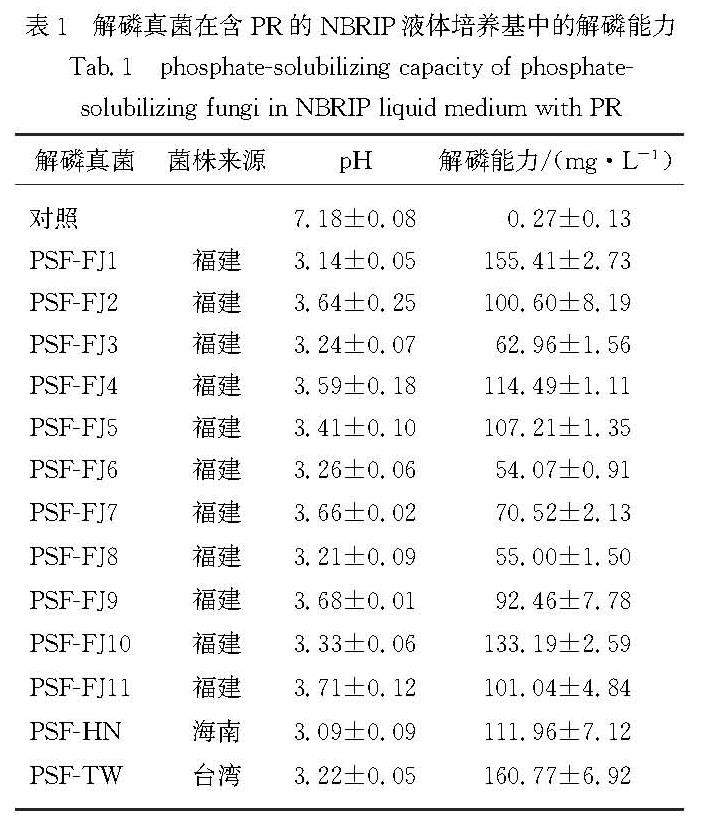
如图1所示,高质量浓度Pb使得多株解磷真菌的菌丝体颜色由白色或浅黄色转为褐色或黑褐色,且大部分真菌菌株在生长后期的孢子形成受到严重抑制.这与Huang等[27]的研究结果类似,在400 mg/L Pb的胁迫环境下,与对照处理相比,木质素降解菌Phanerochaete chrysosporium的菌丝体由最初的规则乳白色变成不规则的黄色短菌丝,真菌菌株可通过改变菌丝密度和菌丝分泌物等对Pb胁迫作出响应.本研究也发现部分菌株在生长后期形成深色分泌物.
本研究采用Jiang等[22]方法中的SLP固体培养基对供试菌株的Pb耐受能力进行分析,尽管已选用低磷含量的培养基,但Pb的加入仍无法避免白色沉淀的产生,该结果表明固体培养基中Pb的实际有效态含量低于预设浓度,因此供试菌株的MIC值将被高估.此外,经过较长时间的培养,部分解磷真菌如PSF-HN(图1(c))菌落中央出现一定程度的溶解现象,说明随着培养基中养分的消耗,部分菌株会重新活化原先被培养介质固定的Pb.
(a~c)解磷真菌PSF-HN:(a)为PDA培养基中菌株的形态,
(b)(正面)和(c)(背面)为Pb质量浓度2 000 mg/L的
SLP培养基中菌株的形态,红色箭头示意透明溶解圈;
(d~f)解磷真菌PSF-FJ11:(d)为PDA培养基中
菌株的形态,(e)(正面)和(f)(背面)为Pb质量
浓度2 000 mg/L的SLP培养基中菌株的形态.
图1 重金属Pb对解磷真菌形态的影响
Fig.1 Influence of heavy metal Pb on morphological characteristics of phosphate-solubilizing fungi
综合考虑供试菌株对PR的溶解能力和对Pb的耐受能力,优选菌株PSF-FJ1、PSF-FJ3、PSF-TW和PSF-HN进行后续研究.基于形态学特征
图2 4株供试解磷真菌在PDA培养基中的形态特征
Fig.2 Morphological characteristics of the four selected phosphate-solubilizing fungi on PDA medium
(图2)和分子鉴定结果,PSF-FJ1、PSF-FJ3、PSF-TW和PSF-HN这4株解磷真菌分别被鉴定为Penicillium sp.PSF-FJ1、Penicillium sp.PSF-TW、Aspergillus niger PSF-FJ3和A. aculeatus PSF-HN.
2.3 液体培养条件下Pb对解磷真菌生长的影响
SLP固体培养基常被用于分析微生物对重金属Pb的耐受能力[22,28-35],但其中的0.05%(质量分数)磷酸根离子和0.15%(质量分数)硫酸根离子会与重金属Pb反应形成白色沉淀物.与重金属总量相比,微生物对游离金属离子的含量更敏感[10].Saeki等[32]的研究也指出,重金属的有效态含量对微生物生长的影响远比重金属总量更重要.因此,本研究参照Deng等[33]的方法尝试使用PDB培养基,但仍无法避免白色沉淀物的产生.
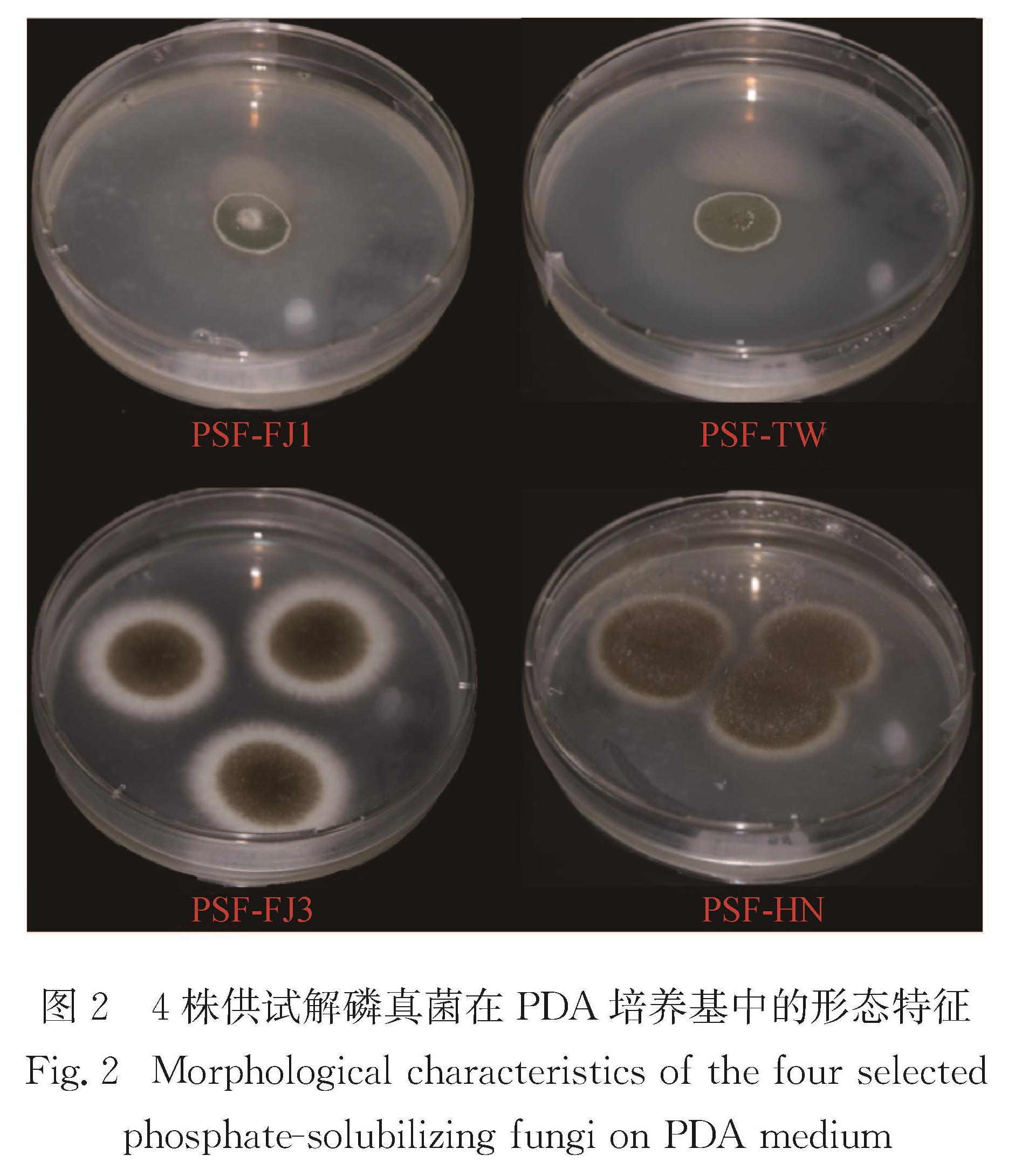
图3 4株供试解磷真菌在不同Pb质量浓度的液体培养基中的耐受系数
Fig.3 Tolerance indexes of the four selected phosphate-solubilizing fungi in liquid medium of different Pb mass concentrations
如图3所示:在200 mg/L Pb的液体培养条件下,PSF-FJ1、PSF-TW、PSF-FJ3和PSF-HN这4株解磷真菌的重金属耐受系数IT均达到0.6以上,其中菌株PSF-FJ3对Pb的耐受系数最高; 当培养液中Pb的质量浓度达到500 mg/L时,菌株PSF-FJ3仍表现出最高的耐受系数,但菌株PSF-FJ1的生长则受到明显抑制; 随着培养液中Pb质量浓度的提高,所有菌株的生长都受到不同程度的抑制.此结果与Park等[10]的研究结果一致,培养液中有效态Pb的实际含量低于初设浓度.同时通过比较可见,菌株在固体培养基和液体培养条件下对Pb的耐受能力有所差异,在Pb质量浓度低于1 000 mg/L的固体培养基中,4株菌株的生长只受轻微抑制作用,菌株甚至在Pb质量浓度高达2 500 mg/L的固体培养基中仍存活较好.
2.4 Pb胁迫抑制解磷真菌的PR溶解能力
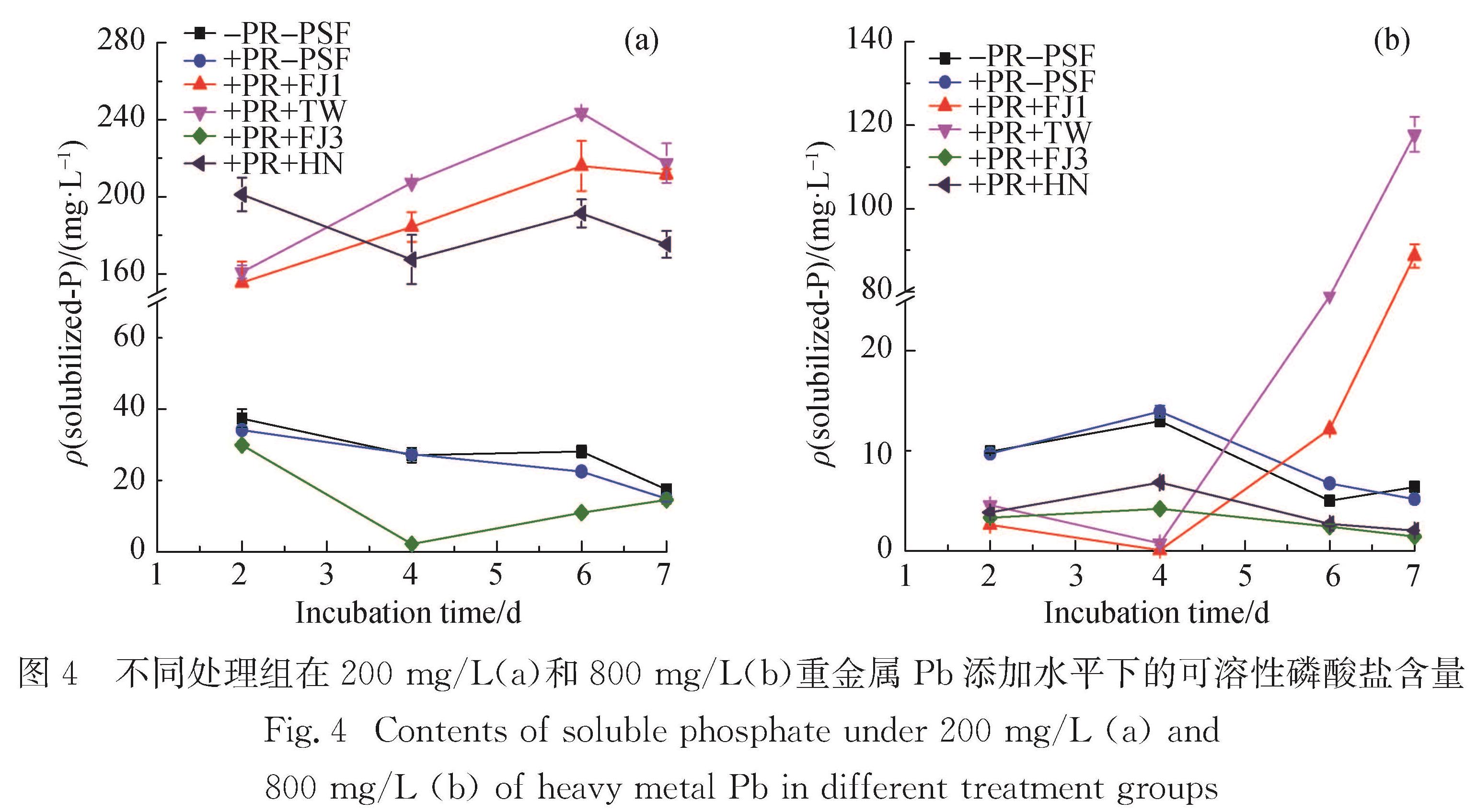
从图4可见,不接种菌剂的对照处理水溶液培养体系(-PR-PSF 和+PR-PSF)中,无论是否添加PR都可检测到一定量的可溶性磷酸盐,且更高浓度的Pb溶液中可溶性磷酸盐的含量会进一步降低.培养介质组分(尤其是磷酸盐)会固定部分游离Pb离子,
图4 不同处理组在200 mg/L(a)和800 mg/L(b)重金属Pb添加水平下的可溶性磷酸盐含量
Fig.4 Contents of soluble phosphate under 200 mg/L(a)and 800 mg/L(b)of heavy metal Pb in different treatment groups
导致Pb的实际含量低于初始设定浓度,如Pb初设质量浓度为200 mg/L时检测到的实际值只有约50 mg/L,Pb初设质量浓度为800 mg/L时检测到的实际值也仅约260 mg/L.
在不同Pb质量浓度下,与不接种处理相比,PSF-FJ3的参与不仅不能提高溶液中可溶性磷酸盐的含量,反而因大量菌丝体的生长消耗了培养体系中的可溶性磷酸盐; 而PSF-HN虽在Pb质量浓度较低(200 mg/L)时可高效溶解PR,但当Pb质量浓度达到800 mg/L时则丧失解磷能力,菌丝体的生长同样消耗了溶液中原有的少量可溶性磷酸盐.由此可见,Pb胁迫会抑制部分解磷真菌的解磷功能.Pérez-Rama等[34]的研究也指出,当培养液中的Cd质量浓度从0.6 mg/L提高到45 mg/L时,菌株Tetraselmis suecica开始呈现重金属Cd的毒害作用.
与菌株PSF-FJ3和PSF-HN不同的是,菌株PSF-FJ1和PSF-TW通过自身代谢活动从PR中释放大量可溶性磷酸根离子,使得溶液中的可溶性磷酸盐含量显著增加; 然而,高质量浓度(800 mg/L)Pb胁迫导致解磷真菌达到最高解磷量所需的培养时间延后,在培养至第6天时,培养液中的可溶性磷酸盐含量才迅速上升.
2.5 耐Pb解磷真菌联合PR有效钝化重金属Pb
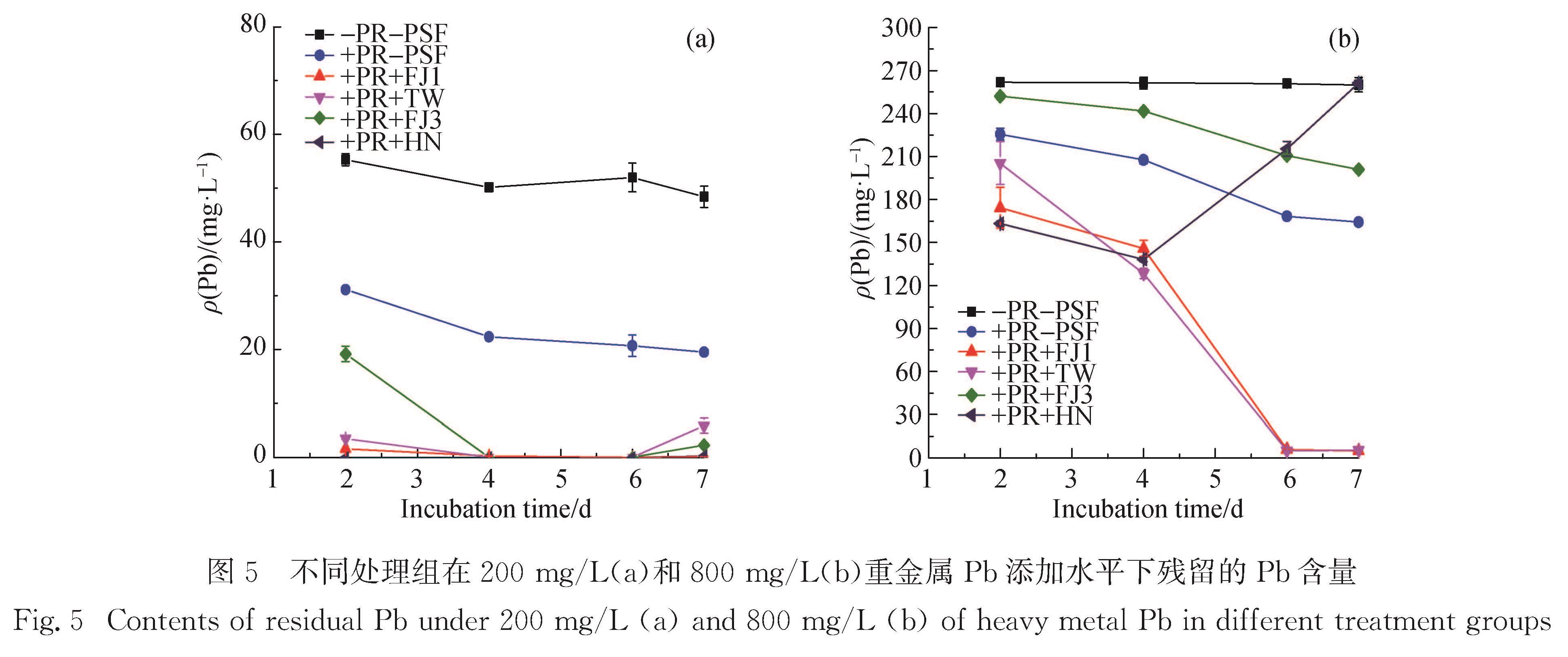
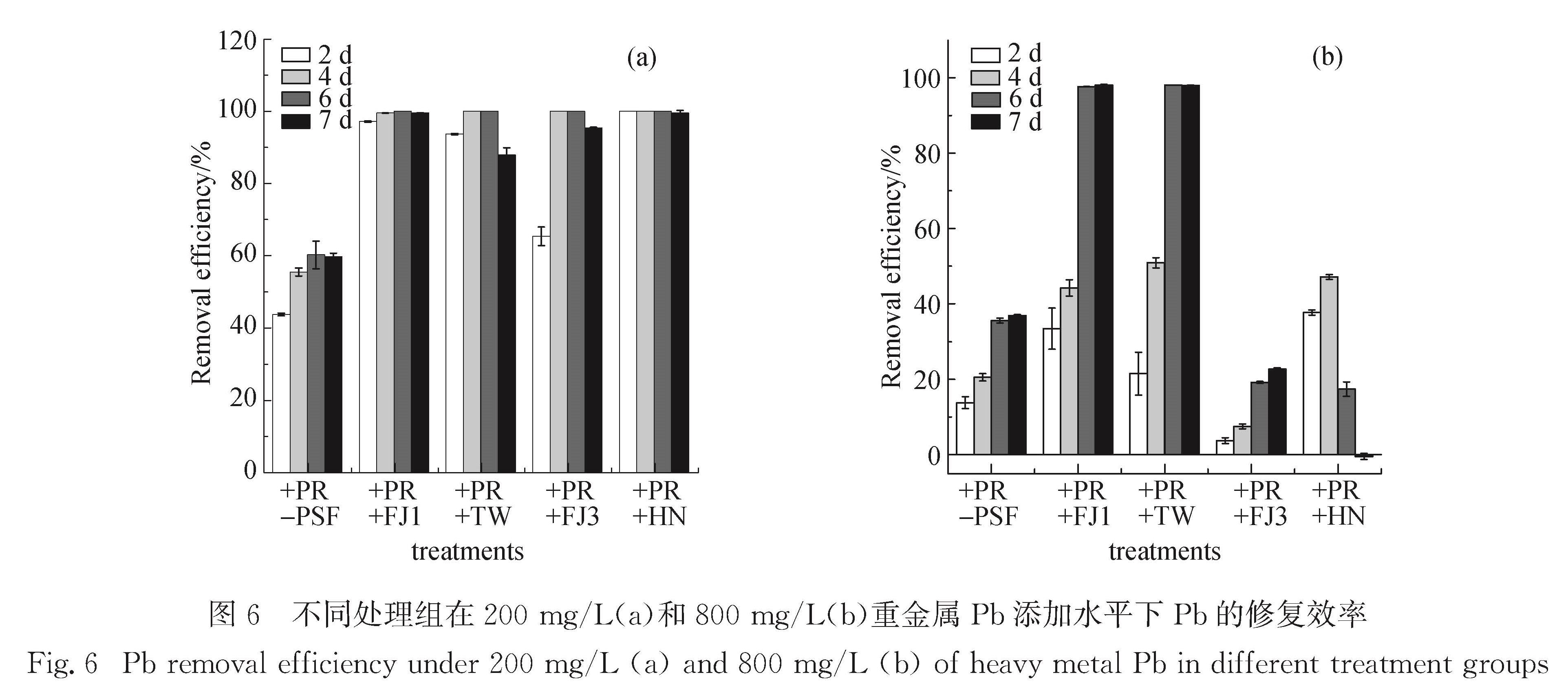
本研究发现PR可在一定程度上降低溶液中Pb离子的含量,但单一PR处理的钝化能力有限,PR的低溶解度会抑制其对重金属的钝化修复效率(图5和图6).如图5(a)和6(a)所示,当Pb质量浓度为200 mg/L时,随着菌株的菌丝体生长和代谢活动,4株菌株的参与都使得溶液中残留Pb含量降至极低水平,重金属Pb得以有效固定,可见解磷真菌在较低水平的胁迫环境下对Pb的钝化能力与其解磷能力不成正比.微生物的吸附作用是其钝化重金属的重要机制,Yang等[35]和Chen等[36]的研究结果显示,一株分离自红树林土壤中的内生真菌菌株Fusarium sp.#ZZF51,通过自身的吸附作用有效钝化重金属U(Ⅵ).从生长在重金属污染土壤的Brassica chinensis根系中分离出的Pb和Cd耐受
图5 不同处理组在200 mg/L(a)和800 mg/L(b)重金属Pb添加水平下残留的Pb含量
Fig.5 Contents of residual Pb under 200 mg/L(a)and 800 mg/L(b)of heavy metal Pb in different treatment groups
图6 不同处理组在200 mg/L(a)和800 mg/L(b)重金属Pb添加水平下Pb的修复效率
Fig.6 Pb removal efficiency under 200 mg/L(a)and 800 mg/L(b)of heavy metal Pb in different treatment groups
内生真菌Mucor sp.CBRF59,可通过菌丝体中细胞膜的运转作用实现Cd和Pb在细胞内的积累[33].此外,Li等[17]借助X射线衍射和扫描电子显微镜-能谱技术,发现菌株A.niger在溶解氟磷灰石时分泌的草酸代谢物可与重金属Pb反应生成草酸铅沉淀,从而有效去除溶液中的Pb离子.在较低污染水平的重金属胁迫环境中,本研究优选的4株解磷真菌可通过菌丝体的生物吸附或代谢产物的分泌有效实现重金属Pb的钝化修复,从而利用功能微生物的大量繁殖实现污染环境的持续修复.
然而,在200 mg/L Pb胁迫条件下,与丧失PR溶解能力的菌株PSF-FJ3相比,保留有解磷能力的其他3株菌株可在更短时间内实现Pb的钝化修复,菌株溶解PR释放的自由磷酸根离子有助于在更短时间内实现钝化修复效果.随着培养时间的延长,PSF-TW、PSF-FJ3和PSF-HN因大量菌丝体生长对营养物质需求的增加,会重新释放原先被固定的重金属Pb,这与长时间培养后菌株在含Pb的SLP固体培养基上菌落中心区域形成明显溶解圈的现象一致.营养物质的大量消耗会加速菌株的死亡[37],有些解磷细菌可将原先反应形成的难溶性含Pb复合物Pb5(PO4)3Cl等作为磷源供自身生长需要[38-39],再次活化重金属Pb,原先吸附在菌丝体的Pb或与有机酸螯合的Pb有被重新释放到溶液中的风险.因此,利用微生物技术钝化修复重金属时,不可忽略修复周期和生长环境中的营养状况等重要因素.
微生物对重金属的修复效率受到重金属浓度的影响,菌株自身对重金属的生物吸附能力有限,如Yan等[40]、Chen等[41]和Deng等[33]的研究发现,菌株对Pb的吸收量在35.7~50.9 mg/g之间.当重金属污染水平超过微生物的吸附能力,为了适应高浓度的重金属胁迫环境,有些菌株通过合成酶或金属结合肽来缓解重金属毒害作用,进一步提高细胞对重金属的吸收能力[33].如图5(b)和6(b)所示:当Pb质量浓度达到800 mg/L时,菌株PSF-HN在培养前期随着菌丝体的生长表现出对Pb浓度的进一步降低效果; 但累积到一定程度后,菌株细胞对有毒金属的外排作用占主导[33].与Park等[16]在液体培养体系中的研究结果相似,在高浓度Pb胁迫环境中丧失解磷能力的曲霉属菌株PSF-FJ3和PSF-HN无法实现对水溶液体系中Pb的钝化修复,反而活化了原先被培养体系或PR钝化的部分Pb离子.
除了传统的非特异性生物吸附作用之外,真菌菌株通过代谢过程介导形成低溶解度的铅矿物沉淀是重金属Pb得以钝化修复的另一种重要机制[1].Sowmya等[42]从印度重金属污染土壤中筛选出的耐U(Ⅵ)解磷细菌(Acinetobacter sp.YU-SS-SB-29),通过代谢分泌的有机酸溶解难溶性磷酸钙盐后,释放出的磷酸根离子可与重金属U(Ⅵ)形成黄色的磷酸铀矿物沉淀.本研究在高质量浓度Pb胁迫环境中,经过6 d培养后,保留PR溶解功能的2株青霉属菌株(Penicillium sp.PSF-FJ1和Penicillium sp.PSF-TW)将溶液中Pb含量迅速降低至极低水平,溶液中残留的Pb质量浓度分别仅为(4.93±0.44)mg/L和(5.37±0.37)mg/L,Pb的钝化修复效率分别达到98.10%和97.93%.两株解磷真菌从PR中释放出的大量可溶性磷酸盐,与溶液中游离的Pb离子发生钝化反应,可高效钝化修复重金属污染的水溶液,实现水体净化.本研究中青霉属菌株的生长导致培养体系的酸化,部分pH甚至降低到4.2以下(结果未展示),但与Park等[10]的研究结果不同,酸化现象并未抑制解磷真菌对重金属Pb的钝化修复能力.该结果可归因于解磷真菌的菌丝生物量远超于解磷细菌的生物量,菌丝体表面有大量吸附位点,且解磷真菌比解磷细菌具有更高的解磷能力,供试菌株供应自由磷酸根离子的能力可实现水溶液体系中重金属的钝化修复.
上述结果表明,解磷真菌对重金属Pb的钝化修复效率与菌种类型、修复周期、生长环境营养状况和重金属污染水平等密切相关.由于滨海湿地高盐、酸性等生境特殊性,从红树根际微区域中筛选出的2株青霉属解磷真菌(Penicillium sp.PSF-FJ1和Penicillium sp.PSF-TW)在联合PR钝化修复重金属Pb污染方面更具优势.同时,这两株青霉属解磷真菌具产生植物生长素和铁载体的潜力(结果未展示),可作为农业有益菌强化植物修复重金属污染的能力.