收稿日期:2019-08-27 录用日期:2020-01-03
基金项目:国家自然科学基金(31870780); 福建省自然科学基金(2019J01021)
通信作者:qwang@xmu.edu.cn
基金项目:国家自然科学基金(31870780); 福建省自然科学基金(2019J01021)
通信作者:qwang@xmu.edu.cn
(厦门大学生命科学学院,福建 厦门 361102)
(School of Life Sciences,Xiamen University,Xiamen 361102,China)
DOI: 10.6043/j.issn.0438-0479.201908026
曲酸是酪氨酸酶的有效抑制剂,然而不稳定性和毒副作用等问题限制了其在美白化妆品中的应用.该研究合成了一种新型的曲酸衍生物KAD3,通过酶学实验、小鼠黑色素瘤B16F10细胞模型实验和斑马鱼模型实验,分析了KAD3对酪氨酸酶的抑制作用以及对黑色素形成的抑制作用机制.结果表明:KAD3是蘑菇酪氨酸酶的混合型可逆抑制剂,能够显著抑制二酚酶活力(半数抑制浓度IC50=10.00 μmol/L); KAD3还能够有效抑制小鼠B16F10细胞内酪氨酸酶活性以及黑色素合成相关蛋白MITF、TYR、TYRP1和TYRP2的表达,随着KAD3浓度的升高,p-Akt表达显著上调,p-CREB 表达明显下调,最终抑制黑色素的形成; 此外,KAD3可以抑制斑马鱼胚胎中黑色素的形成,且效果优于同等浓度的曲酸.上述结果可为曲酸衍生物KAD3在化妆品和医药等领域的应用提供理论参考.
Kojic acid is a well-known anti-tyrosinase agent,however it has been banned to be used in cosmetics due to its instability and side effect.In this study,a novel kojic acid derivative KAD3 was synthesized and the inhibitory mechanism was evaluated with enzyme assay,mouse melanoma B16F10 cell assay and zebrafish assay.The results showed that KAD3 as a mix-type reversible inhibitor of mushroom tyrosinase,could effectively inhibit the diphenolase activity with IC50 of 10.00 μmol/L.KAD3 could effectively inhibit the tyrosinase activity and melanogenesis by downregulating melanin related proteins MITF,TYR,TYRP1 and TYRP2 in the B16F10 cells.p-Akt expression was upregulated while p-CREB expression was downgregulated significantly,with the increasing KAD3 concentration.Moreover,we found that KAD3 could inhibit the melanin formation in zebrafish embryo and the effect was stronger than kojic acid.Our study could supply the theoretical references for application of kojic acid derivatives in the fields of cosmetics and medicine.
随着时代变迁,人们逐渐不满足于化妆品带来的表面暂时性美白,开始追求皮肤自然透出的白皙,因而市面上各种所谓天然美白产品层出不穷[1].目前,广泛使用的美白剂有胎盘提取物、氢醌、熊果苷(对苯二酚葡糖苷)、果酸、曲酸及其衍生物、维生素C及其衍生物等[2].这些物质都已被证实有一定的美白效果,但用于人体时可能出现过敏、脱皮甚至引发疾病.已有研究发现氢醌在淡化肤色的同时也产生了许多副作用,长期使用氢醌还会引发外源性白斑病和褐黄病[3].
黑色素和动物的肤色、毛色以及虹膜的颜色密切相关; 黑素细胞是一种能够特异性产生黑色素的细胞,常见于皮肤、毛囊以及眼睛等部位[4].黑色素能够抵御紫外线从而保护皮肤和眼睛,此外还具有防止内部组织过热的作用[5].哺乳动物的黑色素细胞中酪氨酸酶(TYR)含量丰富,在黑色素合成过程中起关键作用,其异常表达会引发一系列的皮肤疾病[6].若TYR过量表达,会造成色素累积从而导致雀斑、黄褐斑和黑色素瘤等皮肤疾病[7],且这些疾病多发于亚洲女性中[8]; 若TYR表达不足或功能缺失,则会造成眼球、皮肤和毛发的黑色素缺乏而导致白化病[9].此外,TYR家族基因还和某些染色体疾病相关[10].
曲酸因能够有效抑制TYR的活力而闻名,可作为紫外线保护剂,抑制皮肤色素沉积和黑色素形成,过去常作为美白化妆品的重要成分.然而,近些年来有报道称曲酸不稳定、易分解,还有副作用等问题,而被禁止用于化妆品中[11].为此,大量的曲酸衍生物被设计合成,其中大多数是通过改变C-7羟基获得的新型曲酸衍生物,被认为是化妆品、食品、医药等领域中有效的TYR抑制剂.Xie等[12]设计合成了14种曲酸唑类衍生物,对TYR均有明显的抑制作用并且效果显著优于曲酸本身,抑制效果最好的化合物的半数抑制浓度(IC50)达到1.5 μmol/L,比曲酸提高了十几倍; Zhao等[13]以曲酸为母环合成了8种羟基吡啶酮衍生物,并测定了其对TYR活力的抑制作用以及和铜离子结合的能力,结果表明化合物与铜离子鳌合的能力越强则抑制效果越好,并能够有效地延长新鲜切片苹果的货架期.Ashooriha等[14]以曲酸为主要醇基设计了一系列含有芳氧基甲基-1氢-1,2,3-三唑-1-酰基的曲酸衍生物,该类化合物能够有效地抑制蘑菇TYR(mTYR)的活性(IC50=0.06~6.80 μmol/L),显著优于曲酸,特别是两个环烷氧基类似物的抑制作用比曲酸强31~155倍.然而,目前部分曲酸衍生物由于其易被空气氧化、原料不易获取或制备方法复杂等问题而未被商业化使用[15].本课题组前期也尝试对曲酸进行结构改造,先后合成了一系列曲酸衍生物[16].
本研究选取其中一种曲酸衍生物KAD3作为研究对象,从酶学角度探究其对mTYR的抑制作用机制,基于黑色素瘤细胞模型探究其对黑色素形成的抑制效果及抑制机制,并在斑马鱼模型中进一步验证KAD3对斑马鱼(Barchydanio rerio)胚胎黑色素形成的抑制作用,以期为KAD3在美白化妆品和医药方面的应用提供理论依据和实践基础.
人源肝细胞LO2由厦门大学医学院胡天惠教授实验室惠赠; 小鼠黑色素瘤细胞B16F10购自中国科学院上海生物化学与细胞生物学研究所; 斑马鱼胚胎由厦门大学生命科学学院左正宏教授实验室惠赠.mTYR和L-3,4-二羟基苯丙氨酸(L-DOPA)为Sigma-Aldrich公司产品,酶活力为1 000 U/mg; 其他试剂均为国产分析纯试剂; 使用的蒸馏水为去离子双蒸水.黑色素合成相关蛋白MITF、TYR、TYRP1、TYRP2、α-MSH、Akt/p-Akt、CREB/p-CREB等一抗均购自Abcam公司,兔二抗和鼠二抗以及β-actin 抗体购自Abclonal公司.
曲酸衍生物KAD3合成反应过程如图1所示:第一步,
参考文献[13]的方法将曲酸(5 mmol/L)加入30 mL氯化亚砜溶液中,冰浴条件下搅拌反应12 h,反应结束后用二氯甲烷抽滤洗涤沉淀,得到白色粉末状的曲酸氯化物(反应i); 第二步,以4-氨基-3-肼基-5-巯基-1,2,4-三唑(5.5 mmol/L)与肉桂醛(10 mmol/L)为反应物,两者分别用无水乙醇溶解后在玻璃反应瓶中混合,加入少量对甲苯磺酸作为催化剂,反应体系在80 ℃下油浴搅拌加热并冷凝回流,约1.5 h后用无水乙醇抽滤,洗涤沉淀得到席夫碱类产物,为淡黄色粉末(反应ii); 第三步,将第二步合成的席夫碱(3 mmol/L)溶解在10 mL二甲基甲酰胺(DMF)中并缓慢滴加三乙胺(3.3 mmol/L),然后将第一步合成的曲酸氯化物(3 mmol/L)溶解在5 mL DMF中,并缓慢滴加到上述反应瓶中,80 ℃油浴搅拌加热并冷凝回流,反应 12 h(反应iii).反应结束后,加入大量的双蒸水使沉淀析出,用0.1 mol/L稀盐酸洗涤沉淀3遍,抽滤得到棕色沉淀.最终产物用液相色谱-质谱(Waters2767,Waters公司)、核磁共振(AVANCE-Ⅲ600 MHz,Brucker公司)和红外光谱(Nicolet iS5,Thermo Fisher公司)进行鉴定.
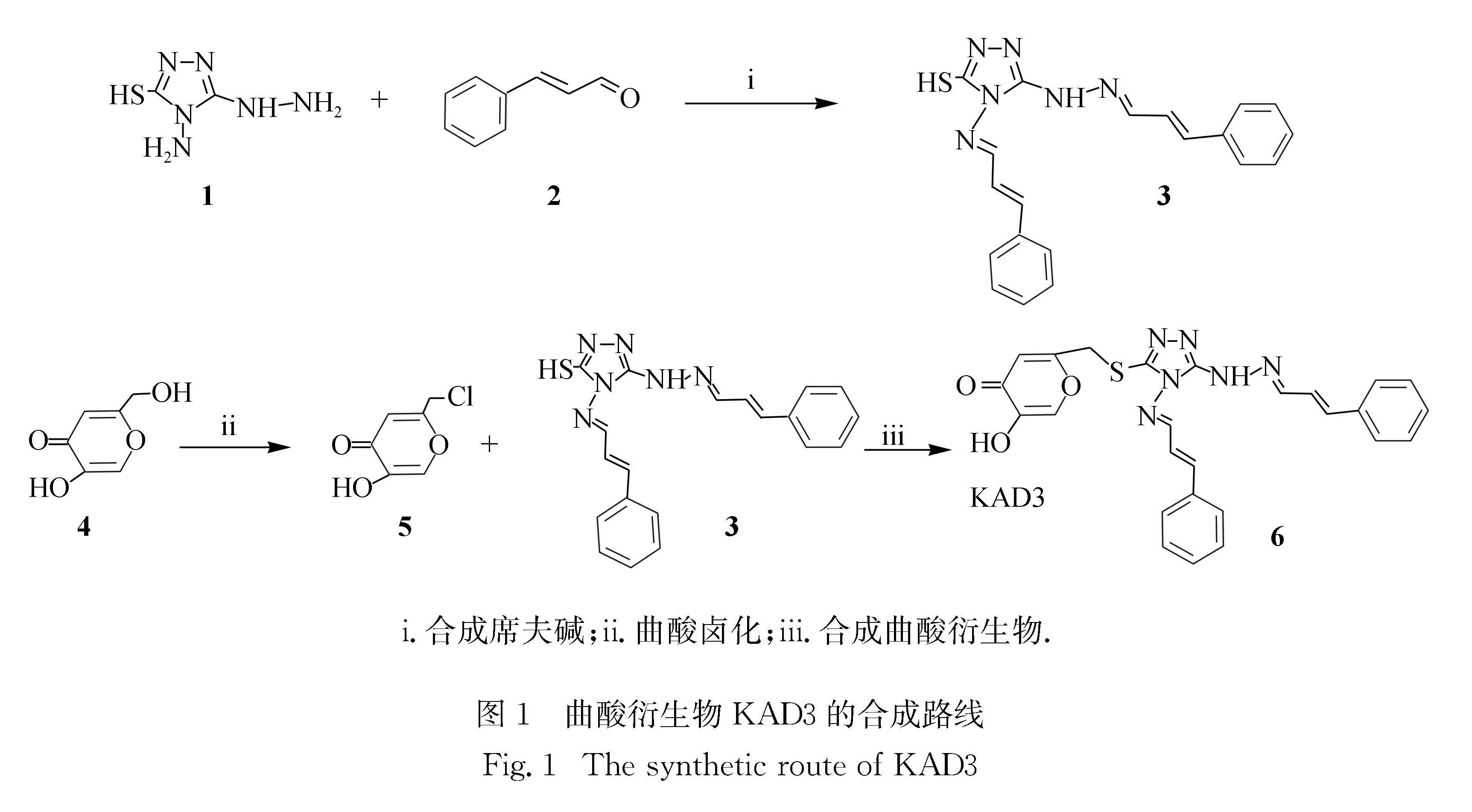
图1 曲酸衍生物KAD3的合成路线
Fig.1 The synthetic route of KAD3
参考文献[17]的方法,以L-DOPA作为底物测定mTYR二酚酶活力.在3 mL反应体系中,依次加入1.65 mL双蒸水、0.75 mL 50 mmol/L磷酸缓冲液(pH 6.8)、0.5 mL L-DOPA溶液、0.1 mL不同浓度的KAD3,最后加入0.1 mL 酶液,迅速混匀后在30 ℃ 恒温条件下测定475 nm处的吸光度.消光系数按照ε=3 700(mol/L·cm)-1计算,直线斜率即为酶活力.以酶的相对剩余活力对抑制剂浓度作图,通过SPSS软件计算效应物的IC50值.
以0.5 mmol/L L-DOPA为底物,通过改变mTYR的浓度,探究不同浓度的KAD3对不同浓度的mTYR催化L-DOPA氧化的影响.以KAD3作用后酶的相对剩余活力对加入的mTYR浓度作图,由此判断效应物对mTYR二酚酶的抑制机制.
在确定KAD3是可逆抑制剂的基础上,进一步探究化合物的抑制类型.在3 mL测活体系内,固定酶浓度而改变L-DOPA的量,检测不同浓度的KAD3影响mTYR催化L-DOPA氧化的酶活力.用双倒数作图法求得抑制常数KI和KIS.
采用Pymol软件(Schrödinger公司)和MOE(2008)软件(Chemical Computing Group公司)将KAD3与TYR进行分子对接,得到TYR-KAD3复合物的模拟结构,从分子水平上分析KAD3与TYR的相互作用机制.从PDB数据库(http:∥www.rcsb.org)下载mTYR(PDB:2Y9W)的晶体结构并去除其外源蛋白、水分子及聚乙二醇,只保留一个含铜亚基作为TYR的蛋白模型.在进行分子对接前,先将蛋白及KAD3分子进行能量最小化,随后选择距离铜离子活性中心最近的区域作为结合位点,进行KAD3最优折叠构象的搜索,根据软件给出的评分标准和KAD3的最终构象,选择最优的复合物结构进行TYR和KAD3的相互作用分析.
采用噻唑兰(MTT)法测定KAD3对LO2细胞和B16F10细胞增殖的影响.用DMEM高糖培养基培养LO2细胞和B16F10细胞,控制细胞培养箱温度37 ℃、5%(体积分数)CO2、饱和湿度的条件下进行培养.待细胞长至融合状态,用0.25%(质量分数)胰蛋白酶消化细胞,并迅速转移至96孔板培养; 每孔加20 μL 单细胞悬液,过夜生长贴壁后吸去原培养基,分别加入0,5,10,20,40,80,100,200 μmol/L的KAD3,每个浓度6个重复,继续培养24或48 h后吸去旧培养液; 于结束前4 h,用pH 7.4的磷酸盐缓冲液(PBS)洗涤细胞3次,每孔细胞加入20 μL的0.5 mg/mL MTT和180 μL新鲜的DMEM高糖培养基,于细胞培养箱中培养细胞4 h后弃去培养基; 再向每孔加入200 μL DMSO,在室温条件下振荡10 min,使蓝紫色结晶甲臜充分溶解,立即用酶标仪(MULTISCAN GO,Thermo公司)测定其在570 nm处的吸光度.根据下式计算:细胞存活率=(A实验组-A空白组)/(A对照组-A空白组)×100%.
细胞培养方法同1.2.6小节,基于MTT实验结果,在不影响其增殖的浓度范围内设置0,8,15,30,60,120 μmol/L 6个浓度,探究KAD3对B16F10细胞内TYR活力以及黑色素合成的影响,以曲酸作为阳性对照.
1)对胞内TYR活力的抑制
参考文献[18]的方法.加药处理60 h后弃去原培养基,用PBS洗涤一次,再用PBS冲洗6孔板,收集细胞于4 mL离心管中,10 000 r/min常温离心后弃上清,保留细胞沉淀; 在细胞沉淀中加入200 μL 0.01 mol/L PBS(含有1 mmol/L 苯甲基磺酰氟(PMSF)和1%(体积分数)TritonX-100)悬浮细胞后,于-80 ℃中冷冻,反复冻融3次,4 ℃、10 000 r/min离心30 min,上清即为粗酶液; 用Bradford试剂盒(上海生物工程有限公司)测定蛋白浓度,并将所有处理组调整至蛋白浓度一致; 取干净的96孔板,在200 μL的酶活力测定体系中加入150 μL溶于PBS(pH 6.8)的L-DOPA,再加入50 μL 提取的粗酶液,暗室37 ℃孵育30 min,测定475 nm 处的吸光度,即视为TYR活力.
2)对胞内黑色素合成的影响.待细胞加药处理60 h后,10 000 r/min常温离心收集细胞沉淀; 用PBS洗涤沉淀3次,10 000 r/min常温离心后弃上清,沉淀加入200 μL 1 mol/L NaOH 于95 ℃水浴30 min,使细胞破碎,色素溶解; 冷却后测得405 nm处的吸光度,即视为黑色素含量.每一浓度处理设3个重复,取平均值.
细胞培养方法同1.2.6小节,设置KAD3的4个浓度分别为0,15,45和60 μmol/L.加药处理72 h后吸去原培养基,用PBS洗涤2次后收集细胞至4 mL离心管中,10 000 r/min常温离心后弃上清,用PBS洗涤3次,收集细胞后加入200 μL含有1 mmol/L PMSF的RIPA裂解液(北京普利莱基因技术有限公司),充分悬浮细胞,放置在冰上裂解1 h.10 000 r/min 4 ℃离心30 min,取上清液用Bradford试剂盒测定蛋白浓度,分装,加入4×上样缓冲液,100 ℃热变性30 min 以保护蛋白不降解.所得样品用Western blot方法分析MITF、TYR、TYRP1、TYRP2、α-MSH、Akt/p-Akt、CREB/p-CREB和β-actin蛋白的表达水平,具体参照文献[19]的方法.
参考文献[20]的方法,采用野生型斑马鱼,水温和pH分别保持在(28.5±0.5)℃和7.5~8.5,每天光照14 h,喂食2次.取胚胎前一晚,按1:2雌雄比挑选健康的野生型斑马鱼,喂饱后放入交配缸中,插入隔板,次日清晨拨开挡板使雌、雄鱼交配; 在受精后30 min 左右收集斑马鱼胚胎,并培养于28 ℃恒温培养箱中,用新鲜的胚胎培养液洗涤3次,去除死胎后转移至培养皿中,再加入新鲜的培养液放置于鱼房培养,在体式镜下挑选发育正常胚胎用于实验; 以6孔板作为实验容器,每孔40个胚胎.
KAD3先溶解于二甲基亚砜(DMSO)中,DMSO终体积分数为0.5%,KAD3质量浓度设置为0,10,20,40和50 μg/mL.药物处理48 h后,取斑马鱼胚胎于体式镜(德国Leica公司,Leica-M165FC)下观察其形态和黑色素分布并拍照.药物处理48 h(受精72 h)后收集胚胎置于-80 ℃处死,用蒸馏水洗涤3次,加入冰浴的含1 mmol/L PMSF的1%(体积分数)Triton X-100,超声处理5 min,10 000 r/min、4 ℃离心30 min,转移上清为粗酶液,用Bradford试剂盒进行蛋白定量; 然后在6孔板中加入150 μL L-DOPA(1 mg/mL)和50 μL粗酶液,暗室37 ℃孵育30 min,测定475 nm处的吸光度,即视为TYR活力; 沉淀用1 mol/L NaOH重悬,于95 ℃金属浴30 min.冷却后测定405 nm处的吸光度,即视为黑色素含量.
数据以平均值±标准差表示; 通过SPSS 19.0 软件对数据进行单因素方差分析(one-way ANOVA),采用邓肯检验法进行两两比较,p<0.05表示差异显著; 用Graphad软件作图.
肉桂醛和3-氨基-4肼基-5-巯基-1,2,4三氮唑(质量比2:1)合成中间产物,再和
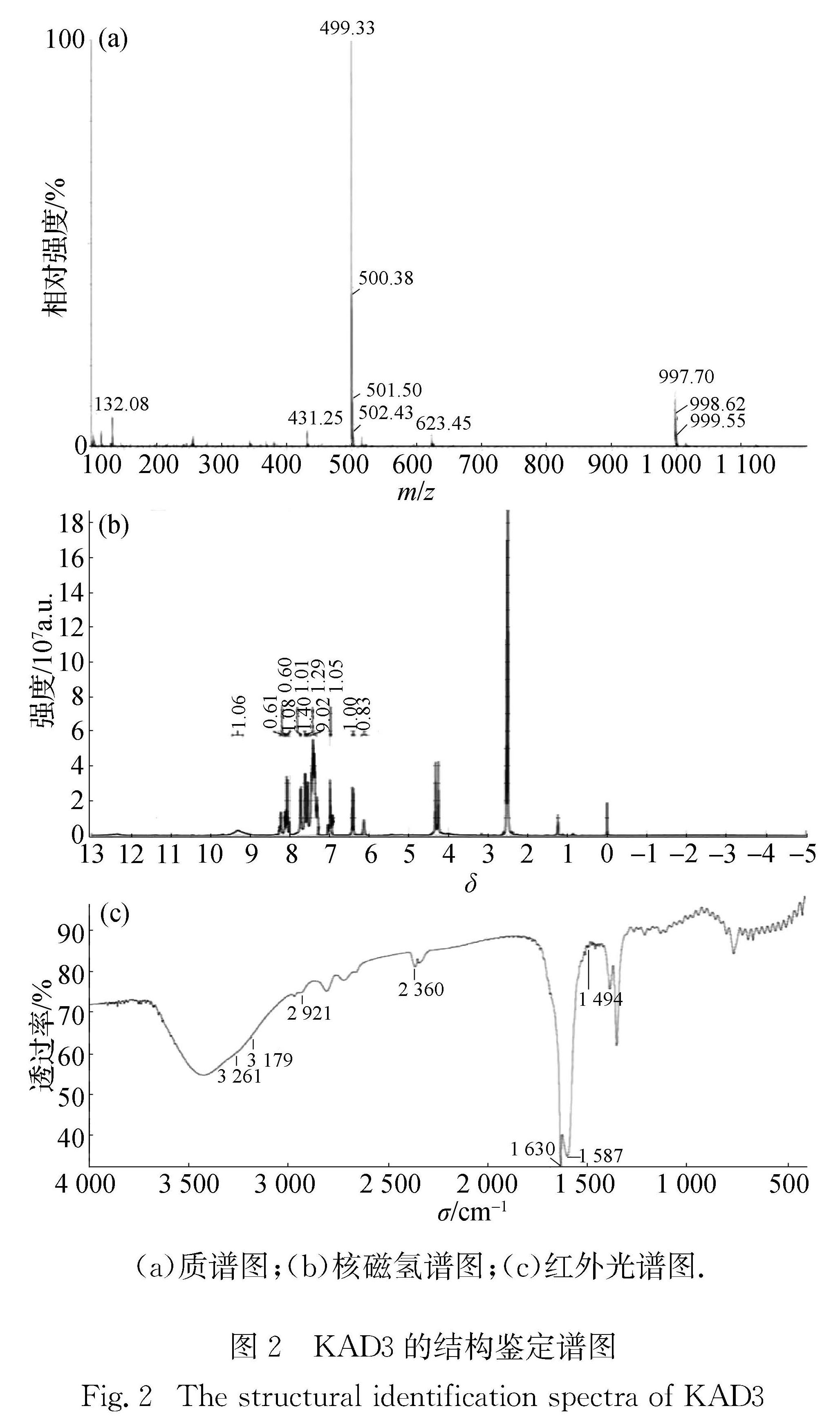
图2 KAD3的结构鉴定谱图
Fig.2 The structural identification spectra of KAD3
曲酸反应得到目标产物KAD3粗产品.经LC-MS提纯(0.035%(质量分数)三氟乙酸水溶液,甲醇梯度洗脱,流速20 mL/min)、旋转蒸发得到棕色固体粉末,产率为60%,纯度超过90%.产物溶于醇类有机溶剂,易溶于甲醇,难溶于水,结构鉴定结果如图2所示.
LC-MS(图2(a))测得相对分子质量为499.33[M+H]+,实际计算值为498.60[M]+.
核磁氢谱(图2(b))数据分析结果如下:1H NMR(600 MHz,DMSO)δ 9.32(s,1H,OH),8.32~8.20(m,1H,pyrone-CH), 8.10(dd,J=31.5,11.4 Hz,2H,N=CH), 7.74(d,J=7.2 Hz,1H,phH), 7.59(dd,J=34.9,7.4 Hz,3H,phH), 7.46~7.34(m,6H,phH), 7.17~7.13(m,1H,N=CH), 7.00~6.96(m,1H,N=CH), 6.92(dd,J=16.1,9.4 Hz,1H,N=CH), 6.45~6.38(m,1H,pyrone-CH), 6.14(s,1H,N=CH), 4.30~4.20(m,2H,S—CH2).
红外光谱(图2(c))数据分析如下(KBr,vmax,cm-1): 3 261, 3 179, 2 921, 2 360, 1 630, 1 587, 1 494.
以L-DOPA为底物,在3 mL反应体系中,研究KAD3对mTYR催化反应的影响,即KAD3对二酚酶活性的抑制作用.结果如图3(a)所示,相对酶活力随着KAD3浓度的增加而逐渐降低.通过SPSS软件计算得KAD3的IC50为10.00 μmol/L,优于曲酸的19.50 μmol/L[21],对二酚酶活性抑制效果显著.通过控制底物浓度不变,只改变酶浓度,测定不同KAD3浓度下二酚酶活力改变情况,进一步研究KAD3抑制mTYR催化反应的机制.结果如图3(b)所示,KAD3浓度越大,直线斜率越小,说明KAD3对二酚酶活性的抑制作用是一个可逆过程,其不能使酶永久失去活性,只能降低酶催化氧化L-DOPA的速率.进一步通过图3(c)-Ⅰ可以看出,1/v对1/c(S)作图得到一组相交于第二象限的直线,说明KAD3 确实能够抑制L-DOPA 的氧化,并且随着Km值的增大Vm值下降,表现为混合型抑制类型; 二次作图,求得KAD3对游离酶的抑制常数(KI)(图3(c)-Ⅱ)为7.50 μmol/L,对酶-底物络合物的抑制常数(KIS)(图3(c)-Ⅲ)为15.80 μmol/L.
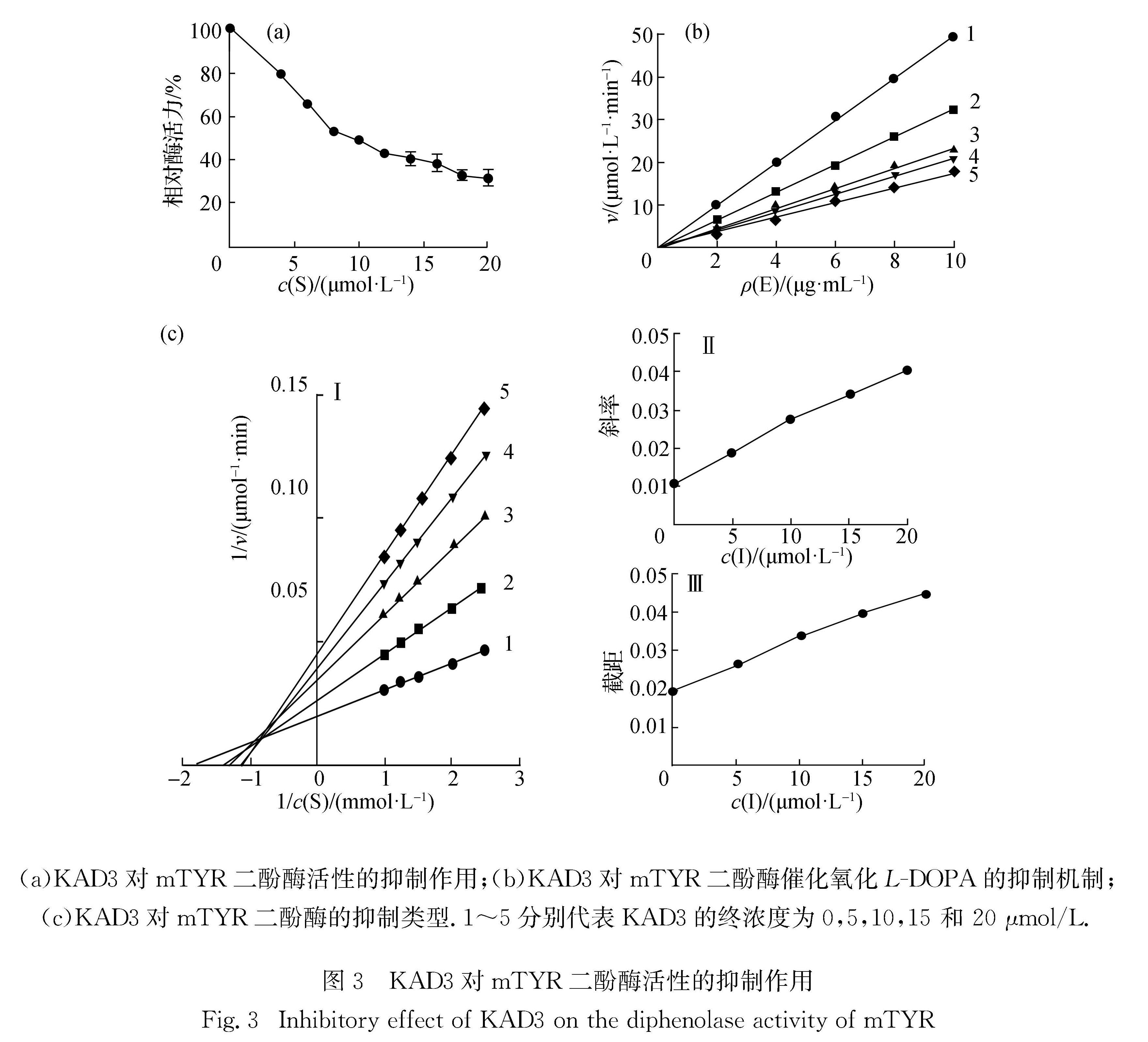
图3 KAD3对mTYR二酚酶活性的抑制作用
Fig.3 Inhibitory effect of KAD3 on the diphenolase activity of mTYR
为了进一步研究KAD3与mTYR的相互作用机制,通过MOE软件进行分子模拟对接,得到mTYR-KAD3的复合物结构,用以分析mTYR和KAD3的相互作用.用Pymol软件和MOE软件作图,mTYR-KAD3复合物的整体结构如图4(a)和(b)所示,KAD3占据了mTYR的催化活性中心(铜离子所
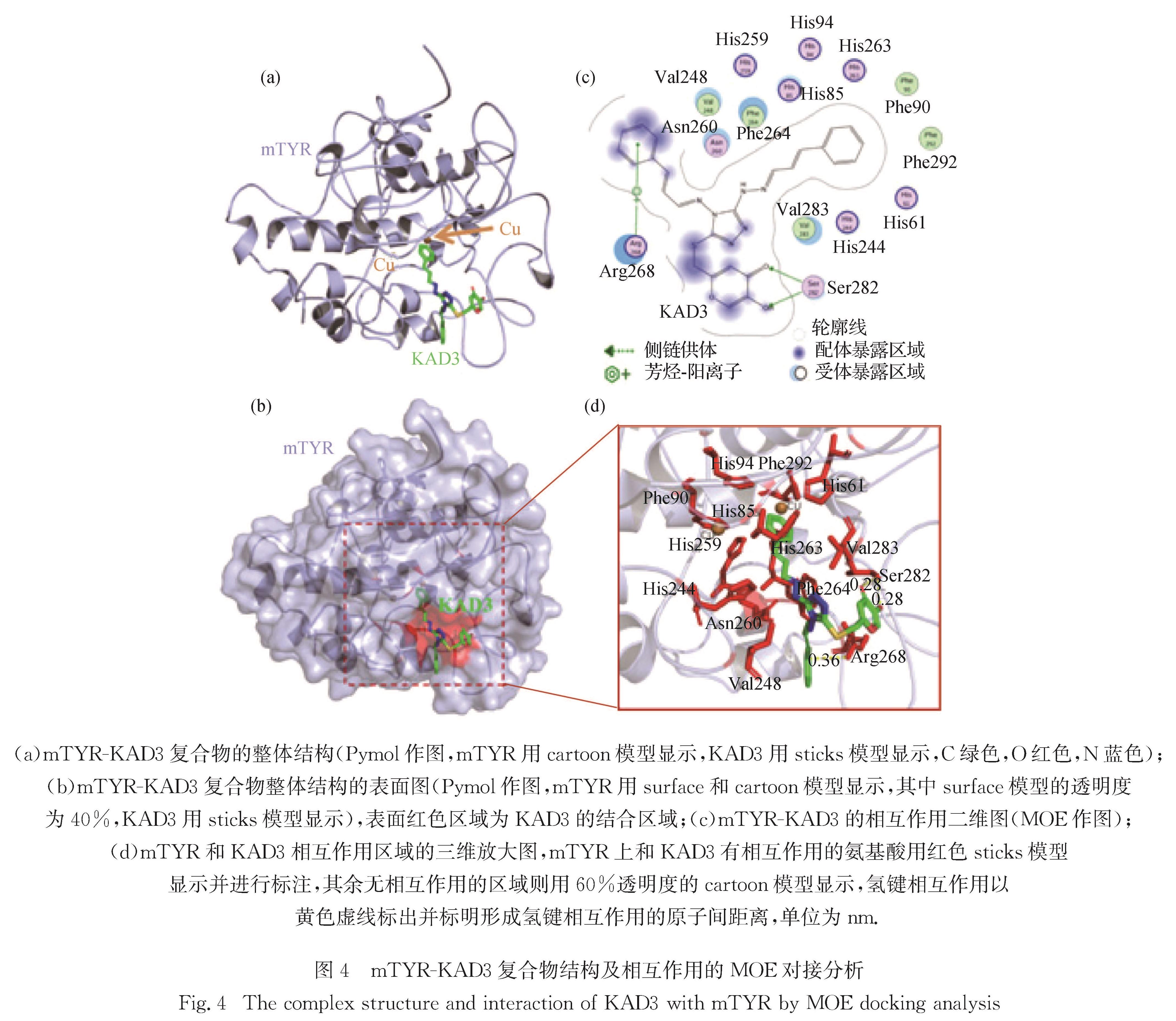
图4 mTYR-KAD3复合物结构及相互作用的MOE对接分析
Fig.4 The complex structure and interaction of KAD3 with mTYR by MOE docking analysis
在的空腔),因此能够抑制mTYR的酶活性.对mTYR和KAD3的结合区域进行相互作用分析,可以看到KAD3和mTYR形成了3个氢键:一个是KAD3的苯基部分与mTYR的Arg268形成的氢键; 另外2个分别是mTYR的Ser282侧链与KAD3酮基上的氧原子和羟基的氧原子形成的氢键.此外,KAD3还与酶活性中心的His259、His263、Ser282、Phe264、Val248、Val283和His94有较弱的相互作用,即范德华力或疏水相互作用(图4(c)和(d)).
利用MTT法检测药物处理后LO2细胞和B16F10细胞的存活率,结果如图5所示:KAD3在0~200 μmol/L范围内对两种细胞的增殖活性均无明显影响,表明在此范围内KAD3对这两种细胞均无毒性,因此下一步实验在此浓度范围内进行.
研究不同浓度(0,8,15,30,60和120 μmol/L)KAD3对B16F10细胞内TYR活力的影响,结果如图6(a)所示:随着KAD3浓度提高,酶活力明显下降,当浓度为120 μmol/L时,酶活力下降至仅剩余2%.从收集的细胞沉淀表观图(图6(b))可以看出,随KAD3浓度提高,细胞的颜色由深变浅,对照组细胞颜色较深而高浓度处理组细胞颜色接近透明.用细胞提取物在405 nm处的吸光度值表征黑色素含量,结果如图6(c)所示:B16F10细胞经过KAD3处理后,黑色素含量显著减少,并且呈现浓度效应,当浓度为120 μmol/L时,黑色素含量仅为对照组的19%.

图5 MTT法检测KAD3对LO2(a)和B16F10(b)细胞增殖能力的影响
Fig.5 Effects of KAD3 on the proliferation ability of LO2(a)and B16F10(b)cells measured by MTT assay

图6 KAD3对B16F10细胞内TYR活力(a)和黑色素含量(b~c)的影响
Fig.6 Effects of KAD3 on the TYR activity(a)and melanin contents(b-c)in B16F10 cells
进一步通过Western blot检测KAD3对B16F10细胞中黑色素合成相关蛋白表达量的影响,从蛋白水平探究KAD3抑制黑色素合成的机制.以0,15,45,60 μmol/L 的KAD3处理B16F10细胞,检测MITF、TYR、TYRP-1、TYRP-2和α-MSH蛋白表达量的差异,以β-actin为内参.从图7可以看出KAD3对MITF、TYR、TYRP-1、TYRP-2和α-MSH的表达均有明显抑制作用.
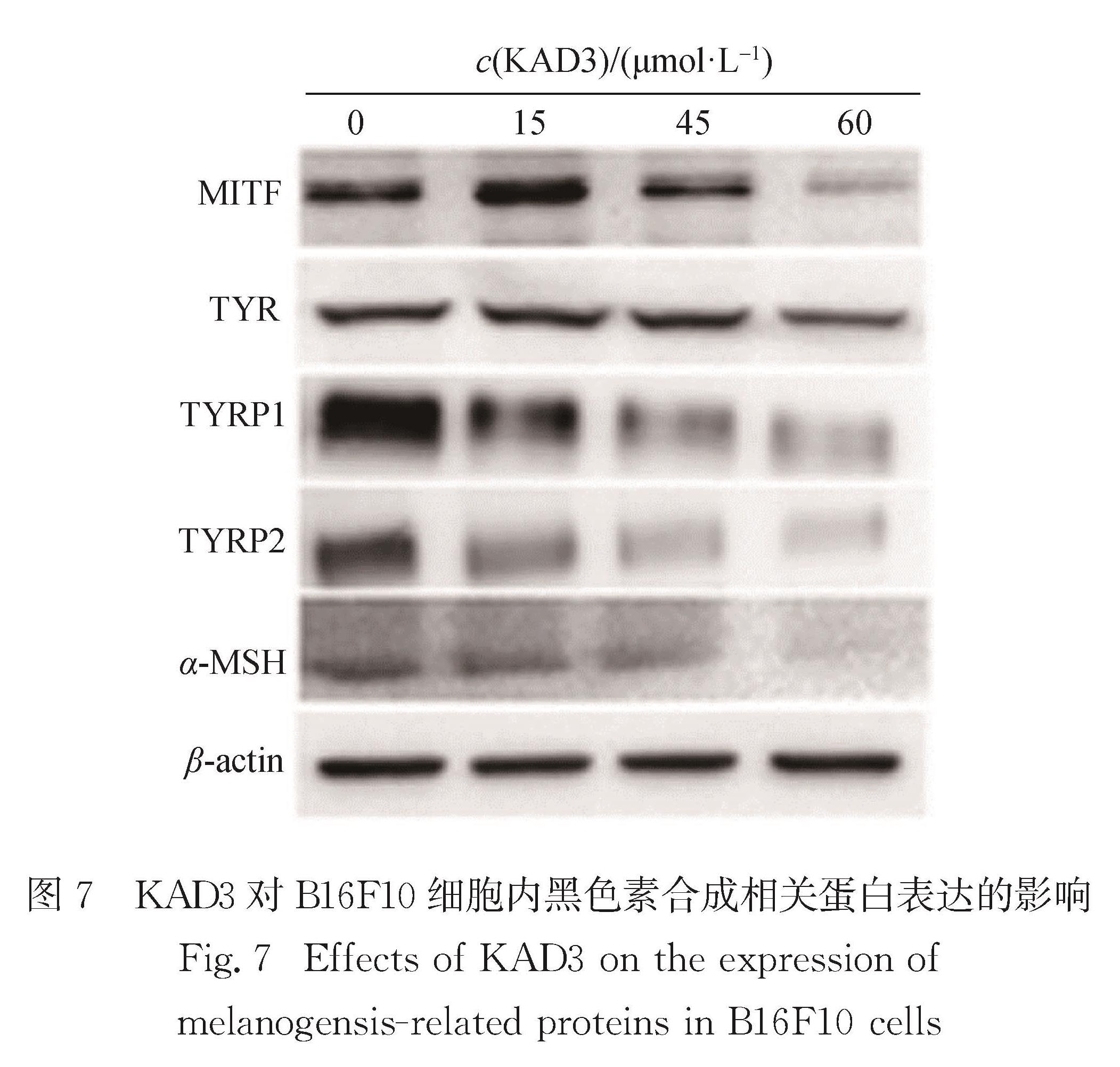
图7 KAD3对B16F10细胞内黑色素合成相关蛋白表达的影响
Fig.7 Effects of KAD3 on the expression of melanogensis-related proteins in B16F10 cells
黑色素的合成是一个复杂的代谢过程,涉及很多信号通路.为探究KAD3对Akt和CREB两条黑色素合成经典信号通路的调控,通过Western blot实验检测Akt/p-Akt、CREB/p-CREB两组蛋白的表达,结果如图8所示:随着抑制剂浓度的升高,p-Akt显著上调,而p-CREB显著下调.

图8 KAD3对B16F10细胞中Akt和CREB信号通路的影响
Fig.8 Effects of KAD3 on the Akt and CREB signaling pathways in B16F10 cells
由于斑马鱼培养简单,黑色素容易观察,常被用来作为筛选黑色素合成抑制剂的动物模型.分别用不同浓度(0,10,20,40,50 μg/mL)KAD3处理斑马鱼胚
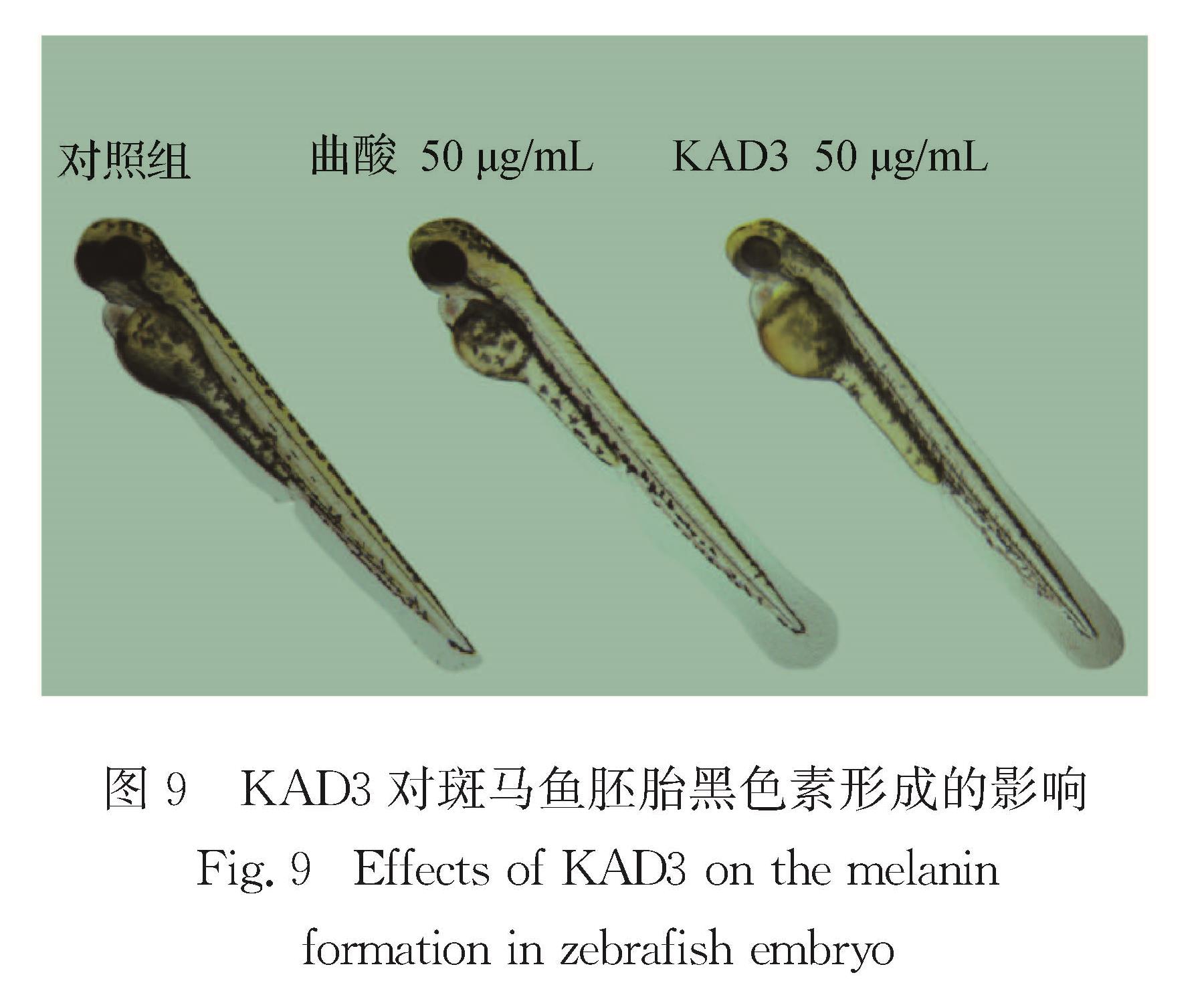
图9 KAD3对斑马鱼胚胎黑色素形成的影响
Fig.9 Effects of KAD3 on the melanin formation in zebrafish embryo
胎,以50 μg/mL曲酸作为阳性对照,观察KAD3对斑马鱼胚胎黑色素形成的影响.如图9所示,50 μg/mL处理组的斑马鱼眼部、卵黄囊的颜色较浅,黑色素形成较少,而对照组颜色很深,并且色素分布密集,可见KAD3处理后斑马鱼胚胎的黑色素形成受到明显抑制,且抑制效果优于同等浓度的曲酸阳性对照组.
进一步的检测结果显示,随着KAD3浓度的增加,斑马鱼胚胎内TYR活力降低(图 10(a))且黑色素含量减少(图 10(b)).
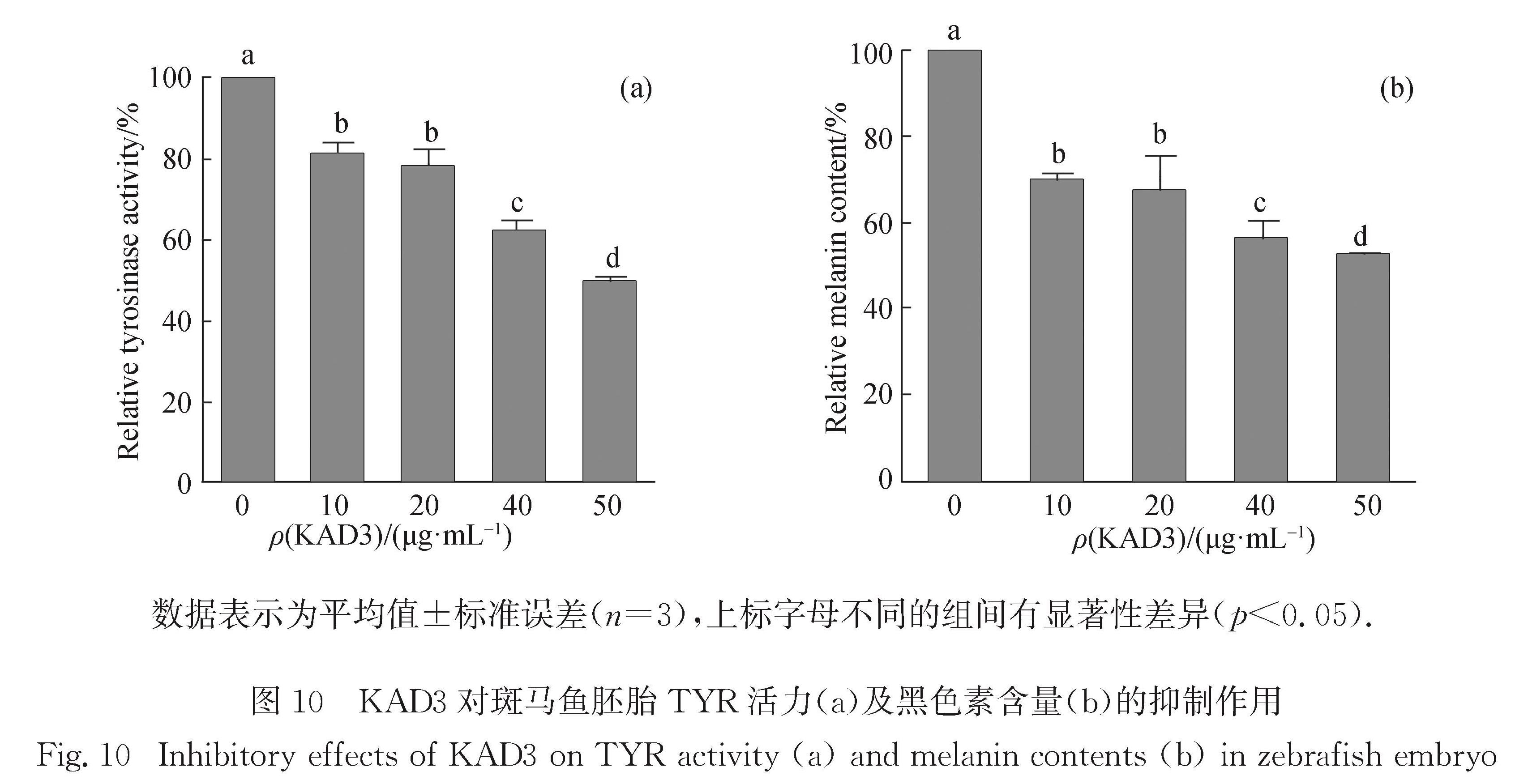
图 10 KAD3对斑马鱼胚胎TYR活力(a)及黑色素含量(b)的抑制作用
Fig.10 Inhibitory effects of KAD3 on TYR activity(a)and melanin contents(b)in zebrafish embryo
本研究以肉桂醛和3-氨基-4肼基-5-巯基-1,2,4三氮唑(质量比2:1)为原料合成中间产物席夫碱,再和曲酸反应得到目标产物KAD3粗产品.经LC-MS提纯、旋转蒸发得到棕色固体粉末.此法相较于其他曲酸衍生物的制备方法更简单易行,且产物不易被氧化.本研究结果表明化合物KAD3对mTYR二酚酶具有显著的抑制效果,并且优于曲酸.该化合物的抑制机制表现为可逆混合型抑制,而且KIS值大于KI,说明酶-底物复合物与抑制剂的亲和力比游离酶与抑制剂的亲和力弱.
MOE分子模拟对接实验结果表明,KAD3可以与mTYR活性位点的部分残基相互作用.侧链供体Ser282可以作用于酮基的氧原子和羟基的氧原子,且酮基的氧原子可以与残基Ser282和Val283形成2个氢键; 此外,KAD3还与活性中心的His259、His263、Ser282、Phe264、Val248、Val283和His94残基相互作用,从而抑制酶的活性.
进一步分析KAD3对B16F10细胞内黑色素合成的抑制作用以及分子机制发现,在没有毒副作用的浓度范围内,该化合物表现出很好的抑制黑色素合成的活性.在120 μmol/L处理下,B16F10细胞内TYR活力几乎被完全抑制,仅为对照组的2%; 而细胞从表观上几乎看不见黑色,以吸光度值表征的黑色素含量仅为对照组的19%.黑色素含量取决于TYR的表达和活性[22].TYR家族相关蛋白TYRP1、TYRP2以及TYR的表达受到关键转录因子MITF的调控,MITF能够与启动子的M-box结合,是细胞黏附分子表达的直接和正向调控因子,并随后促进黑色素合成蛋白的转录[23].本研究结果显示KAD3能够有效下调MITF的表达,进而下调TYR家族蛋白的表达,有效抑制TYR的活性,最终抑制黑色素的合成.为了进一步探究曲酸衍生物抑制黑色素合成的分子机制,本研究还分析了KAD3对黑色素合成的两条主要相关信号通路的调控.有报道表明,cAMP/PKA和MAPK信号通路激活可上调CREB磷酸化,而CREB磷酸化正调控MITF,PKA可磷酸化和激活CREB,进而上调MITF的表达[24].此外,PI3K/Akt信号通路也与黑色素合成相关,失活的PI3K和Akt通过cAMP诱导激活GSK3β,然后在Ser289位点磷酸化MITF,促进其与启动子的M-box结合,增强黑色素原相关酶的表达[25].本研究发现,KAD3可上调细胞中p-Akt 和下调p-CREB的表达,进而下调MITF的表达,最终抑制黑色素的合成.
斑马鱼是一种流行的脊椎动物模型,其器官系统和基因序列与人类相似[26].斑马鱼胚胎黑色素合成是在受精24 h后,其累积肉眼可见,易于观察,因此很多研究者利用斑马鱼体外筛选抑制剂.Adamantia等[27]通过斑马鱼体外表型试验从山楂中鉴定出新的抗黑变抑制剂.本研究中,与对照组相比,在表型上可以明显看出KAD3能够有效抑制斑马鱼黑色素的累积,特别是眼睛和背部的颜色都比较浅,黑色素呈点状分布.
综上所述,本研究结果可为KAD3在化妆品和医药方面的应用提供理论参考.
