自然来源的营养素能有效消除癌症细胞[11],探索天然、高效和低毒的天然药物进行长期的辅助抗癌具有重要意义[12].白凤菜具有多种药理作用,含有丰富有治疗作用的黄酮,适合患者长期食用.有研究证实,高浓度白凤菜水提口服液可阻止小鼠胃肠在机体糖负荷情况下对葡萄糖的吸收[13]; 白凤菜叶提取物能在一定程度上提高抗氧化酶活性和抑制促炎性因子表达,抑制大鼠的炎症[14]; 白凤菜醇提取物对斑马鱼慢性酒精性脂肪肝具有防治作用,能够有效减轻肝脏的炎性病理[15]; 白凤菜醇提取物还可治疗大鼠非酒精性脂肪肝,明显降低非酒精性脂肪肝模型大鼠血脂(甘油三酯和总胆固醇)、丙氨酸氨基转移酶及天冬氨酸氨基转移酶的水平[16]; 白凤菜的乙酸乙酯提取物通过自噬作用抑制HeLa细胞的增殖,而非通过细胞凋亡途径抑制癌细胞增殖[17].本研究通过MTT实验、细胞形态显微镜观察、划痕实验和Annexin V-FITC/PI双染流式细胞术,发现TFG对肝癌HepG2细胞具有抑制增殖和迁移及促进凋亡的作用.
各类细胞在生命周期中不断产生和清除细胞内新陈代谢产物ROS,不同细胞中ROS的表达水平也不同.肿瘤细胞中高水平的ROS有助于激活原癌基因,有利于维持细胞生长和增殖,导致DNA损伤和遗传不稳定性; 同时,肿瘤细胞会提高抗氧化蛋白的表达来消除高ROS的影响,建立氧化还原平衡,维持原癌基因信号通路,防止细胞凋亡[18].有研究证实通过化学或生物学的抗氧化剂降低肿瘤细胞的ROS水平,重新构建细胞内还原系统,可抑制肿瘤细胞的增殖[11,18-19].本研究结果显示当肝癌HepG2细胞被TFG作用后,细胞内ROS水平显著降低(p<0.05),细胞内氧化还原状态改变,可能使细胞在低ROS水平下的相关信号传导途径发生改变,从而抑制肝癌HepG2细胞的生长,并进一步引起细胞凋亡.
Bcl-2和Bax是Bcl-2家族的重要蛋白,与凋亡关系最密切.促凋亡蛋白Bax位于细胞线粒体膜上,寡聚化形成一个跨膜通道,从而增加膜通透性,导致膜电位降低[20].位于内质网膜上的Bcl-2可能通过阻断Ca2+从内质网向胞质的流动,与Bax结合形成异源二聚体,阻止细胞凋亡[21].本研究中65和130 μg/mL TFG实验组Bax的表达量略微增加,Bcl-2的表达量明显降低,而当Bax较Bcl-2相对增高时则可能形成大量的Bax-Bax同源二聚体,两种凋亡调控蛋白间的平衡被打破,进而促使细胞凋亡[22-24].
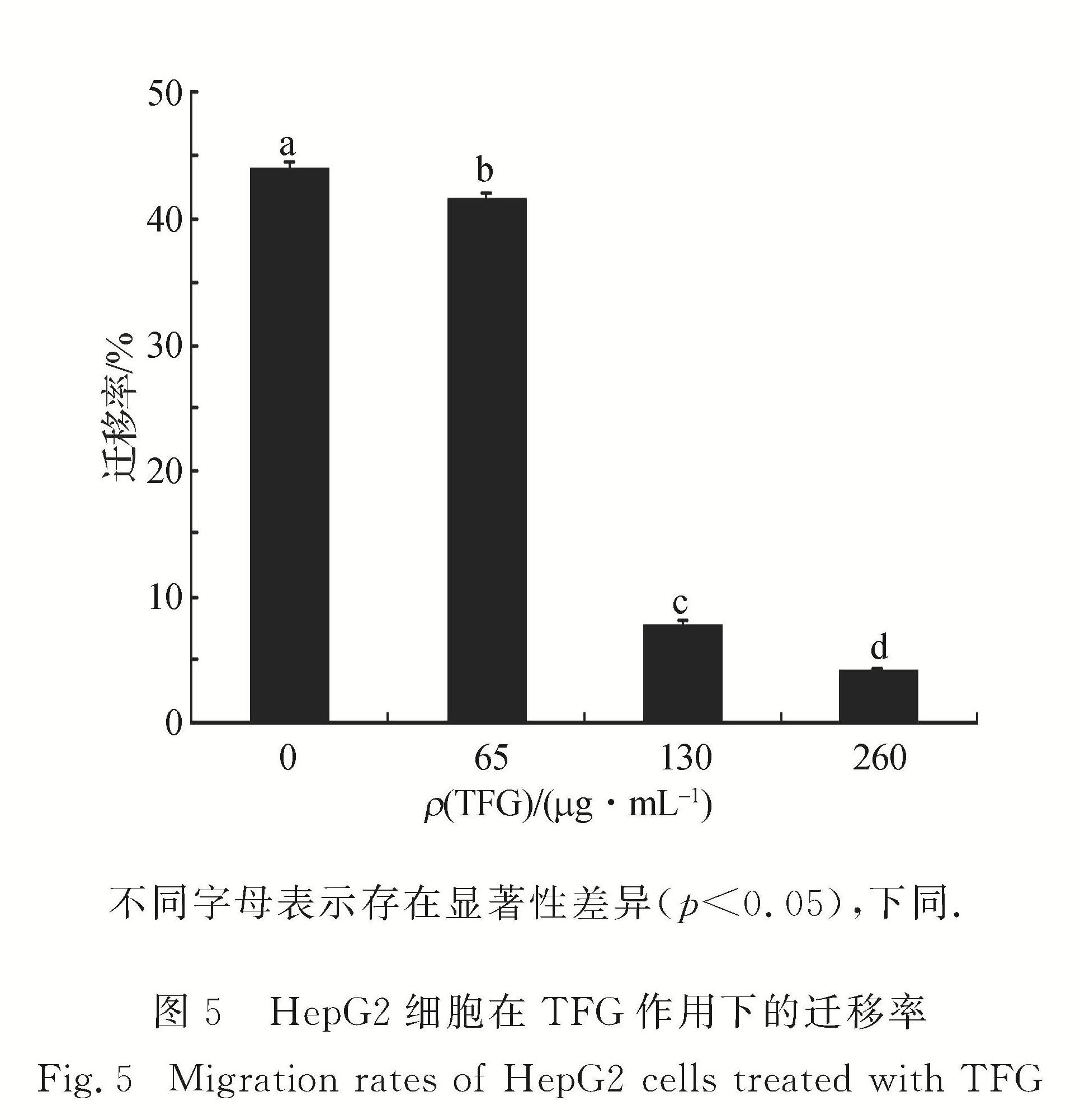
本研究显示不同剂量的TFG作用于肝癌HepG2细胞24和48 h后,当质量浓度小于50 μg/mL时,延长给药时间,细胞存活率无显著性差异; 而当质量浓度大于100 μg/mL时,延长给药时间,细胞存活率有显著性差异,随TFG质量浓度的增加而下降,细胞皱缩,染色质凝集,细胞迁移能力下降,细胞周期被阻滞在S期,荧光凋亡探针呈现阳性,细胞内ROS水平显著降低,凋亡相关蛋白Bax表达量略微上升,Bcl-2表达量明显要下降.综上可见,一定剂量的TFG可抑制细胞活性并诱导细胞凋亡,为进一步研究和开发TFG作为抗肝癌的天然药物提供了理论基础.