2.1 咖啡酸吗啉胺的合成路径及结构鉴定
咖啡酸吗啉胺的合成路径如图1所示,纯化后得到黄色粉末,进行结构鉴定,结果如下:
C-1的IR鉴定(KBr,νmax,cm-1):3 431.28(—OH),1 652.72(C=O),1 276.90(C—N),1 113.07(C—O),968.38(C=C).1H-NMR鉴定(600 MHz,DMSO):δ 8.04(t,J=5.7 Hz,1H),7.23(s,1H),7.20(s,1H),6.96(t,J=4.0 Hz,1H),6.85(d,J=2.1 Hz,1H),6.83(d,J=2.0 Hz,1H),6.77(d,J=3.0 Hz,1H),6.76(s,1H),6.74(s,1H),6.37(s,1H),6.34(s,1H),2.90(d,J=3.3 Hz,1H),2.89(d,J=4.2 Hz,1H),2.88(s,1H),2.64(s,1H),2.55(s,5H).LC-MS鉴定(m/z):测定值293.26 [M+H]+; 理论值292.35 [M]+.
C-2的IR鉴定(KBr,νmax,cm-1):3 473.65(—OH),1 653.73(C=O),1 271.73(C—N),1 118.87(C—O),980.00(C=C).1H-NMR鉴定(600 MHz,DMSO):δ 8.06(t,J=5.6 Hz,1H),7.23(s,1H),7.20(s,1H),6.95(d,J=2.0 Hz,1H),6.87(d,J=2.0 Hz,1H),6.85(d,J=2.0 Hz,1H),6.83(d,J=2.0 Hz,1H),6.75(s,1H),6.74(s,1H),6.33(s,1H),6.31(s,1H),3.18~3.16(m,2H),2.51(dt,J=3.6,1.8 Hz,2H),2.48(s,1H),2.36~2.31(m,5H),2.28(d,J=7.4 Hz,2H).LC-MS鉴定(m/z):测定值307.63 [M+H]+; 理论值306.37 [M]+.
2.2 化合物对酪氨酸酶的激活作用
(a)C-1和C-2作用下的酪氨酸酶活力测定;(b)和(c)分别为C-1和C-2的Lineweaver-Burk双倒数作图,其中曲线0~4对应的
C-1浓度依次为0,0.125,0.250,0.375,0.500 mmol/L,C-2浓度依次为0,0.025,0.050,0.075,0.100 mmol/L.
图2 咖啡酸衍生物对酪氨酸酶的激活作用以及激活类型
Fig.2 Activation activity of caffeic acid derivatives on tyrosinase activity and determination of the activation type of tyrosinase
从图2(a)可以看出,C-1和C-2对酪氨酸酶均具有明显的激活作用,其半激活质量浓度(EC50)分别为0.06和0.12 mmol/L,由此可见,C-1对酪氨酸酶的激活效果约为C-2的2倍.由图2(b)和(c)可知,Lineweaver-Burk双倒数作图均为相交于第二象限的一组直线,直线纵轴截距和直线斜率随着化合物浓度增大而减小,因此可以判断C-1和C-2对酪氨酸酶的激活类型均属于混合型激活.
(a)~(c)分别为体系中未加化合物、加入0.05 mmol/L C-1和C-2在连续时间内L-DOPA氧化产物的累积光谱图,其中
曲线0表示化合物在该波长范围内的光谱,曲线1~11分别代表加入酪氨酸酶0~10 min后的光谱.(d)和(e)分别为不同浓度的
C-1和C-2在相同时间(10 min)内L-DOPA氧化产物的累积光谱图,其中曲线0~6对应的C-1加入浓度依次为0,0.075,
0.150,0.225,0.300,0.375和0.450 mmol/L,C-2加入浓度依次为0,0.015,0.030,0.045,0.060,0.075和0.090 mmol/L.
图3 咖啡酸衍生物对酪氨酸酶催化氧化作用的全波长扫描分析
Fig.3 Full wavelength scanning analysis of caffeic acid derivatives on tyrosinase catalytic oxidation
图3(a)是L-DOPA在未加化合物的情况下,酪氨酸酶催化氧化作用不同时间后的紫外-可见光谱,475 nm处特征峰的累积量为0.41.由图3(b)和(c)可以看出,在体系中添加C-1和C-2,10 min后产物累积量分别增加了78%和58%.由图3(d)和(e)可以看出,随着C-1和C-2浓度的增加,475 nm处的吸收峰也在不断地增加.该结果进一步佐证了2种化合物对酪氨酸酶的激活作用.
2.3 化合物与酪氨酸酶的相互作用
从图4(a)和(b)可以看出,酪氨酸酶的荧光强度随着C-1和C-2浓度的增大而有规律地降低,且荧光发射波长出现略微红移.以F0/F对C-1和C-2终浓度c(Q)作图,得到化合物对酪氨酸酶荧光强度的Stern-Volmer曲线,如图4(c)和(d)所示.通过计算可知,C-1和C-2的荧光猝灭速率常数Ksv分别为7.06×10-6 和1.31×10-5 L/mol; 其动态猝灭速率常数Kq分别为706和13.1 L/(mol·s),均远小于各类猝灭剂对生物大分子的最大Kq值2.0×1010 L/(mol·s),说明C-1和C-2对酪氨酸酶的荧光猝灭均属于动态猝灭机制.
(a)和(b)分别为C-1和C-2作用下酪氨酸酶的荧光谱图,(a)中曲线0~10对应的C-1加入浓度依次为0,0.01,0.02,0.03,
0.04,0.05,0.06,0.07,0.08,0.09,0.10 μmol/L,(b)中曲线0~8对应的C-2加入浓度依次为 0,0.02,0.04,0.06,
0.08,0.10,0.12,0.14,0.16 μmol/L;(c)和(d)分别为不同浓度的C-1和C-2对酪氨酸酶荧光强度的Stern-Volmer曲线.
图4 不同浓度的咖啡酸衍生物对酪氨酸酶荧光强度的影响
Fig.4 Impacts on changes of tyrosinase fluorescence by different concentrations of caffeic acid derivatives
通过分子对接模拟进一步探索了C-1、C-2与酪氨酸酶的对接模式以及酶催化部位,受体所暴露的差异性可通过与氨基酸残基结合的强度显示.从图5的对接构象可以看出,2种化合物均不能与酪氨酸酶的金属铜离子相互作用,但可与活性中心的氨基酸残基相互作用.如图5(a)所示:C-1氨基上的自由氢可以与酪氨酸酶活性中心的Asn260残基形成氢键,杂环供氧可以与Asn260和His244残基形成氢键,这些结构能够改变蛋白质的构象; 此外,C-1还能与活性中心的Val248、Val283、His61、His85、His263、His259、Phe90、Phe264和Glu256残基相互作用.如图5(b)所示:C-2氨基上的自由氢可以与酪氨酸酶活性中心的Asn260残基形成氢键,杂环供氧可以与His61残基形成氢键,这些结构能够改变蛋白质的构象; 此外,C-2还能与活性中心的Val283、His94、His85、His263、His259、Phe264、Phe292、Phe90、Ala286、Arg268、Ser282、Thr84和Glu256残基相互作用.
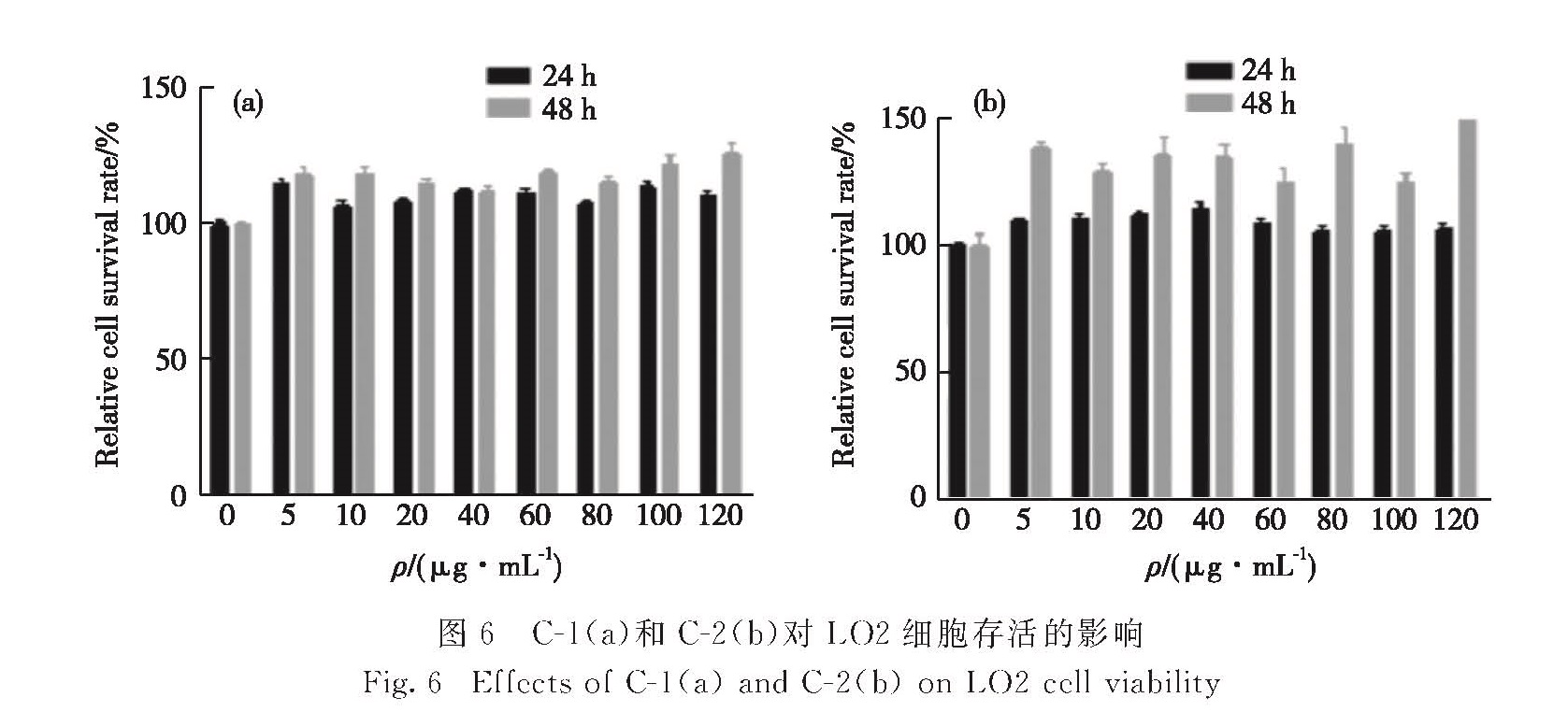
2.4 化合物对人正常肝细胞LO2的毒理试验
通过MTT法检测化合物对人体正常肝细胞LO2的影响.如图6所示,LO2细胞在经过不同质量浓度(5,10,20,40,60,80,100,120 μg/mL)的C1和C2处理24 h后,处理组与对照组之间并无明显差异,而处理48 h后对LO2细胞的存活有一定促进作用,说明C-1和C-2无细胞毒性.
图5 C-1(a)和C-2(b)与酪氨酸酶的氨基酸残基之间的对接模型
Fig.5 Binding modes of C-1(a)and C-2(b)with amino acid residues of tyrosinase
2.5 化合物对M14细胞内酪氨酸酶的激活作用
由图7可以看出,在浓度为0,0.1,0.2,0.3,0.4和0.5 mmol/L 的C-1和C-2的作用下,M14细胞内酪氨酸酶活力增加,且激活作用呈现出浓度依赖性.相同浓度下,C-2对酪氨酸酶的激活效果比C-1好.
图6 C-1(a)和C-2(b)对LO2细胞存活的影响
Fig.6 Effects of C-1(a)and C-2(b)on LO2 cell viability
图7 C-1和C-2对M14细胞内酪氨酸酶活力的影响
Fig.7 Effects of C-1 and C-2 on intracellular tyrosinase activity of M14 cells
2.6 化合物对黑色素合成蛋白表达量的影响
通过蛋白免疫印迹法检测化合物对M14细胞中黑色素合成相关蛋白表达量的影响.由图8可以看出,化合物C-1和C-2对TYR、TRP-1、TRP-2和α-MSH蛋白的表达量均起上调作用,而内参GAPDH蛋白的表达量无明显变化.