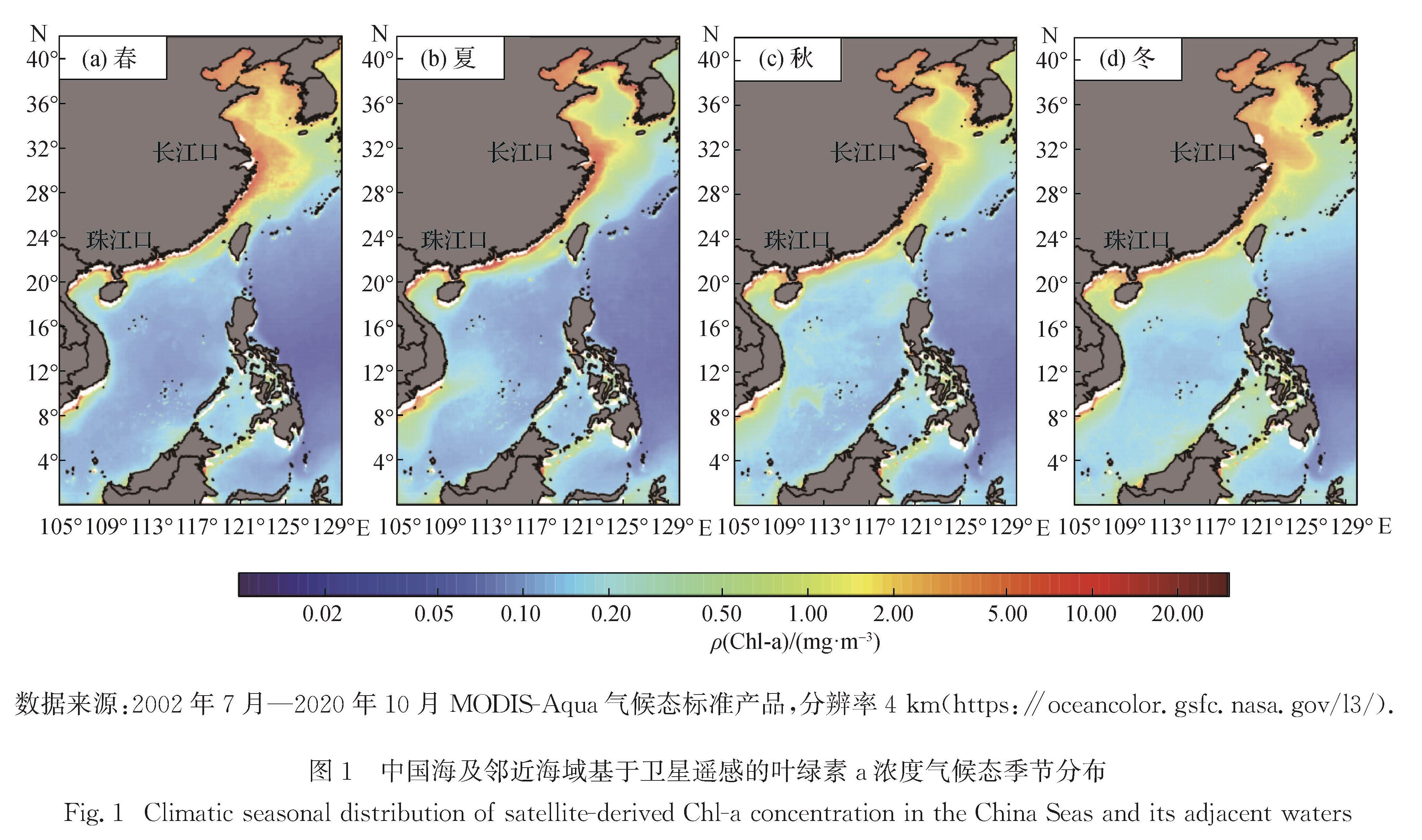
伴随北半球光照强度和日照周期的季节变化和东亚季风的影响,中国边缘海各区域月平均海表光照强度、温度和风速的季节变化规律总体上相似,受此影响,浮游植物叶绿素a质量浓度在季节尺度总体上呈现春、秋季高于冬、夏季的基本分布模式(图1).然而,不同区域因各自独特的地形条件和水文环境,形成了各种不同的水团和海流体系[9].浮游植物群落响应环境变化也因此呈现不同的区域特征和独特的环境调节机制,主要是由光照、营养盐、温度和风速变化驱动的一系列物理-化学-生物耦合过程机制; 与此同时,不同的浮游植物类群对关键环境和生物因子(光照、营养盐、温度和浮游动物摄食等)变化的响应存在显著差异,造成了它们的实际生态位分化和群落时空分布演变,也因此驱动了整个浮游生态系统功能的改变[16-18].
数据来源:2002年7月—2020年10月MODIS-Aqua气候态标准产品,分辨率4 km(https:∥oceancolor.gsfc.nasa.gov/l3/).
图1 中国海及邻近海域基于卫星遥感的叶绿素a浓度气候态季节分布
Fig.1 Climatic seasonal distribution of satellite-derived Chl-a concentration in the China Seas and its adjacent waters
1.1 季节尺度上分布差异的影响因素
春季藻华过程在长江口近岸最明显,叶绿素a质量浓度可达10 mg/m3(图1(a)); 秋季在这些区域藻华现象也时有发生,但与春季相比,浮游植物生物量极值及其范围都显著减小(图1(c)).同样是春季藻华,其强度在不同区域和不同时间均有差异:东海与黄海海域相比,春季藻华发生较晚; 每年冬季至翌年春季期间,各海区叶绿素a质量浓度由北至南依次降低[19].春季藻华主要是温度、光照、营养盐和浮游动物摄食等因素在不同时空尺度上耦合作用的结果,通常呈现不同藻种相互交替的模式[20-21].近年来,部分海区绿藻、金藻和蓝藻藻华也呈上升趋势[22].
夏季受河口羽流(如长江口和珠江口)和上升流(如粤东、浙闽和琼东沿岸)影响,部分近海区域叶绿素a浓度显著提升,与开放海域形成极显著的空间差异(图1(b)):东海夏季长江口冲淡水羽流中叶绿素a质量浓度很高,但跨过盐度锋面陆架表层水中的叶绿素a质量浓度快速下降[23]; 而在黄海中部海区,独特的环流体系使其物理环境相对稳定,形成冷水团和垂直层化结构,造成真光层内浮游植物生物量较低[24-25].黄海冷水团盛行时,中部海区硅藻的叶绿素a质量浓度和相对丰度分别只有0.05 μg/L和30%,而蓝藻的相对丰度为(36±16)%,显著高于其他季节,并在表层水体中占优势地位; 10月垂直层化和黄海冷水团减弱,真光层内定鞭藻的相对丰度较夏季增加[24].基于浮游植物类群水平群落组成的聚类分析结果及温度、盐度、浮游动物(桡足类)甚至鱼类的分布,都表现出一致的空间特征,表明黄海冷水团的理化参数与浮游植物群落及高营养级生物的分布之间都有很密切的关系[24].
与夏季垂直层化结构控制真光层内营养盐供应进而决定浮游植物群落的调控机制不同,冬季叶绿素a质量浓度的相对低值(图1(d))主要是因为光照和温度限制了浮游植物生长; 但南海北部等寡营养海区则与此不同,冬季风带来的垂直混合促进营养盐的向上输送仍然是影响浮游植物群落的主要机制[17,26-28].Xiao等[17]报道南海表层平均总叶绿素a质量浓度在冬季最高,达(0.71±0.76)mg/m3,夏、秋和春季依次为(0.55±0.75)mg/m3,(0.40±0.47)mg/m3和(0.20±0.24)mg/m3.另外,冬季黑潮起源于高温、高盐的黄海暖流和台湾暖流等,在冬末春初风速下降而光照和温度提升的季节变化背景下,与近岸流相互作用进一步提供了有利的水文环境,促进春季浮游植物生长,有利于近岸春季藻华的发生[29].
以上研究结果表明,相对于开放大洋,边缘海生态系统环境复杂,其浮游植物生物量、生产力和群落组成在不同时空尺度下的变化规律,进一步限制了研究者们揭示其调控机制和预测其变化趋势.
1.2 基于生态位特征的时空分布格局
近年来,大量现场实测数据与卫星遥感资料的结合、生态位模型和生态位特征分析等手段的应用,为准确揭示浮游植物时空分布格局和环境调控机制提供了新思路.Liu等[16]整合了在东海7年(2006—2012年)11个现场航次的1 500多个浮游植物样本,结合多源卫星遥感资料,揭示了浮游植物生物量和类群水平群落结构的准气候态季节分布格局,厘清了不同季节浮游植物与3类水团(冲淡水、陆架混合水、黑潮水)相关联的群落组成特征:1)春季模式,表现为冲淡水羽流区形成藻华,甲藻和硅藻的生物量较高; 2)夏季和初秋模式,具有显著的空间差异,表现为冲淡水羽流区高硅藻生物量以及黑潮水中低硅藻生物量和微微型浮游植物占优势; 3)晚秋和冬季模式,生物量相对较低,分布较均匀,但隐藻、青绿藻和金藻的贡献较大.按照这一思路,Xiao等[17]通过整合在南海2004—2015年20个航次的光合色素数据,发现浮游植物的总体分布格局呈现5种类型:1)近岸模式,其特征为南海北部近岸海区全年以硅藻占绝对优势,并伴随有一定浓度的甲藻(春季)、聚球藻和青绿藻; 2)外海模式,其特征为原绿球藻在整个南海的外海水体都具有较高生物量,春夏季最为明显; 3)表层模式,其特征为聚球藻在整个南海的上混合层生物量都较高,但从春季到冬季逐渐减少; 4)次表层模式,其特征为定鞭藻8型和青绿藻在南海次表层的生物量较高,其中定鞭藻8型的生物量最大层比青绿藻的更深; 5)陆架模式,其特征为南海陆架海区浮游植物群落结构具有高多样性,且季节变化较明显.
在现场实测分析的基础上,进一步利用典型相关分析和广义相加模型(GAMs)等机器学习方法,建立主要浮游植物类群对温度、盐度和营养盐等环境因子变化的生态位曲线,并通过生态位曲线的特征分析定量解析主要浮游植物类群的分布特征.Xiao等[17]结合GAMs和最大熵模型(MaxEnt),拟合浮游植物各类群在每个环境因子轴上的实际生态位曲线,并计算每条生态位曲线的平均生态位和生态位宽度,以表征浮游植物对环境变化响应的实际生态位特性,发现南海浮游植物群落结构的变化是各类群在温度、光照和营养盐浓度之间权衡的结果.以此为基础,Zhong等[18]集成了在台湾海峡15年(2004—2018年)25个航次的现场数据和21年(1997—2018年)的水色遥感数据,分析发现:硅藻、青绿藻和绿藻的温度、盐度和营养盐生态位较宽,主要分布在东北季风时期的沿岸水以及西南季风时期的上升流水和冲淡水; 聚球藻和定鞭藻具有相对高温、高盐和中等营养盐的生态位,在中营养海域生物量较高; 隐藻和甲藻因其低温、低盐和高营养盐生态位,在东北季风期间沿岸水较活跃; 原绿球藻具有高温、高盐和低营养生态位,在黑潮影响水体中其生物量较高.因此,结合物种分布模型和GAMs计算浮游植物的生态位,可以通过浮游植物的实际性状解释不同季风期间台湾海峡浮游植物群落的时空分布格局.此外,季风与流场受海峡“窄管”效应影响,造成台湾海峡与其他陆架边缘海相比具有更明显的浮游植物群落时空变异性.
![表1 中国边缘海浮游植物群落的时空格局与演变趋势<br/>Tab.1 !![Spatial-temporal distributions and successional patterns of phytoplankton communities in the Chinese marginal seas]](2021年02期/pic132.jpg)
