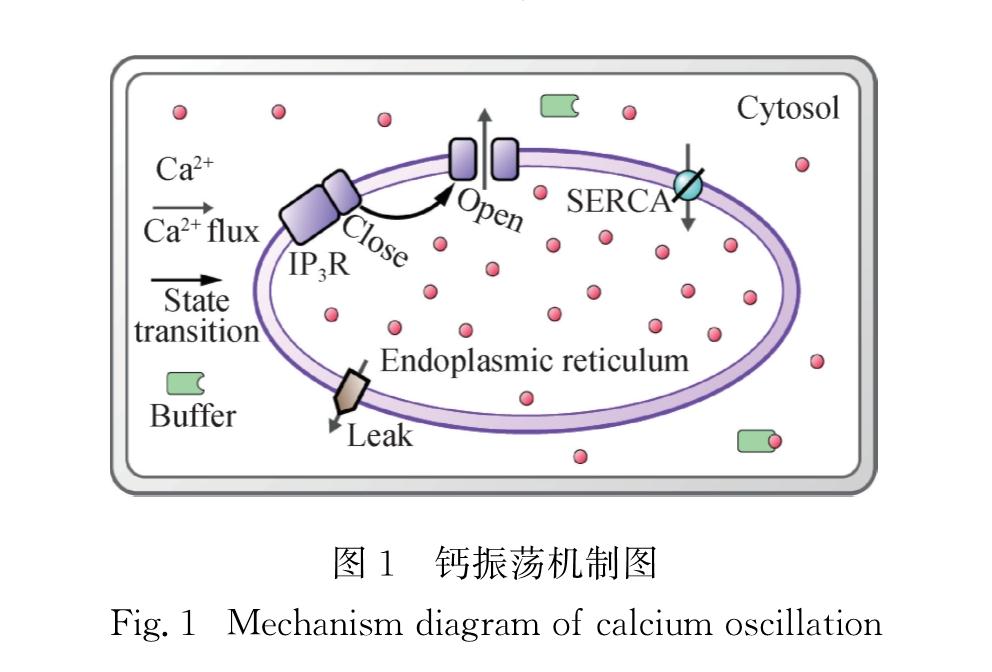
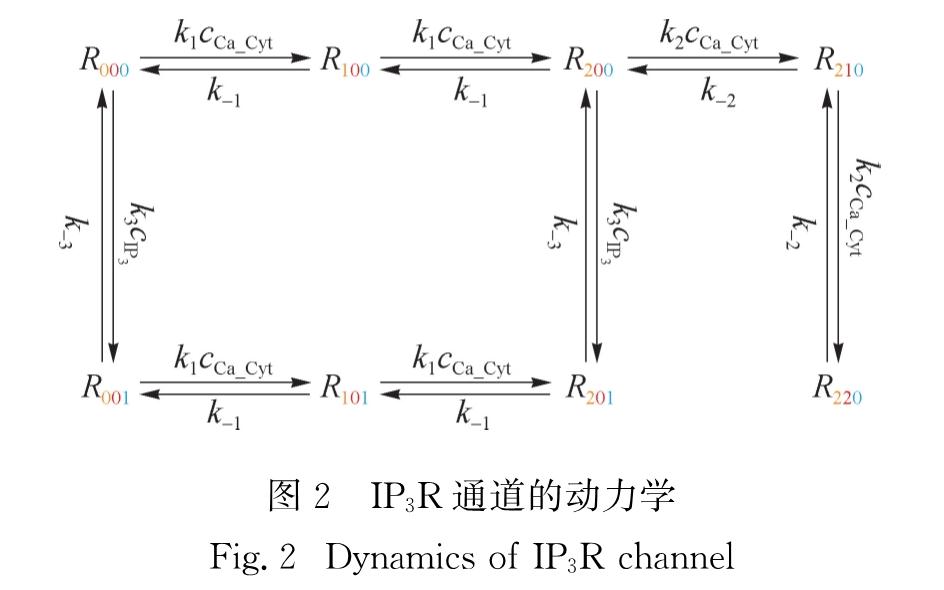
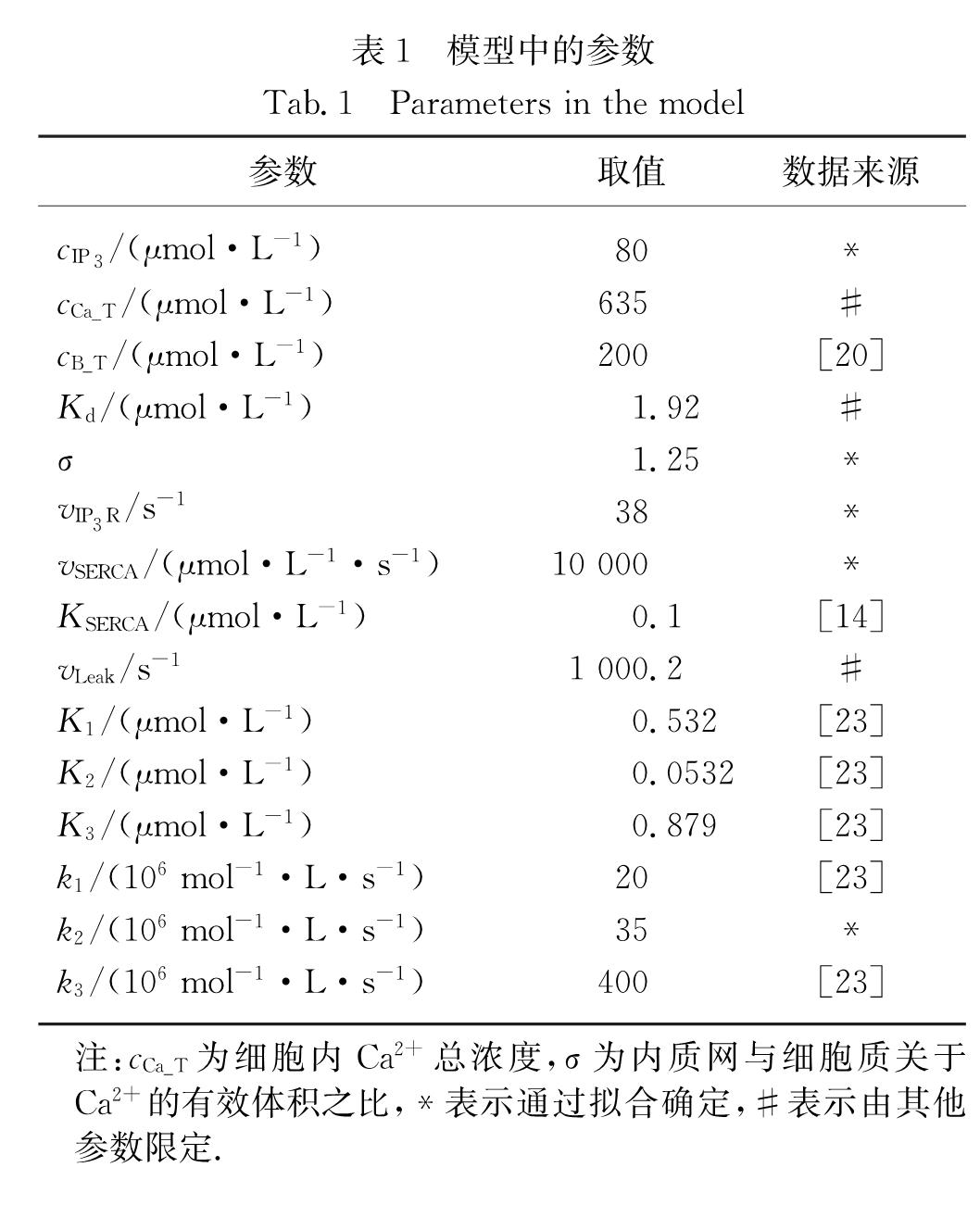
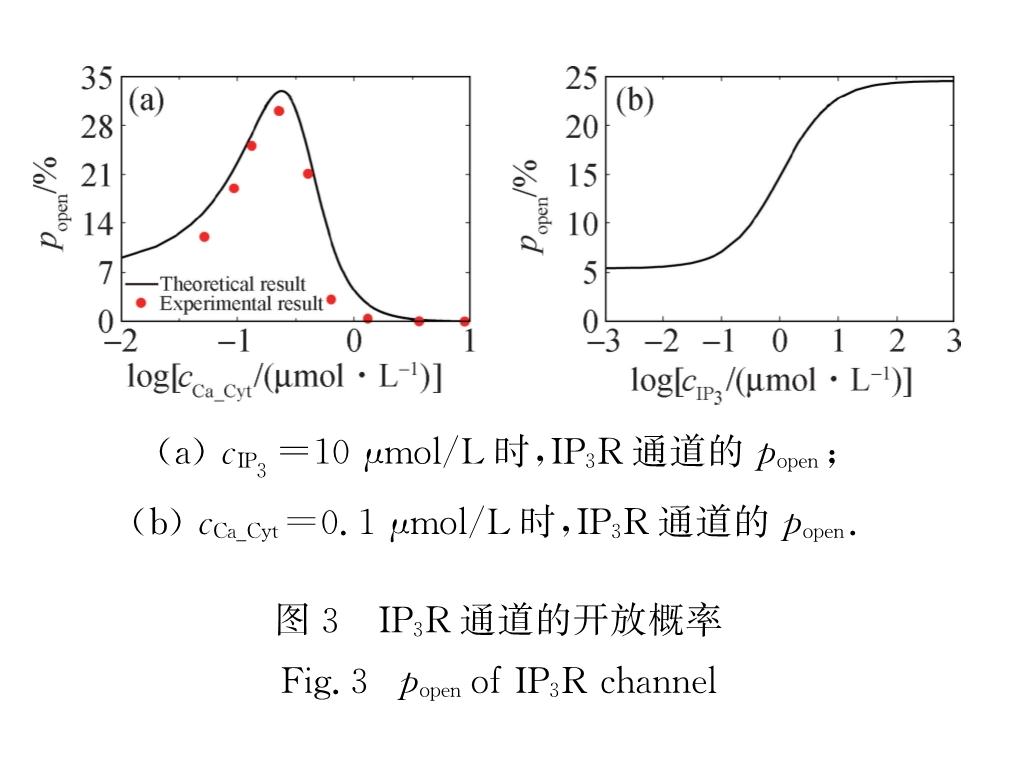
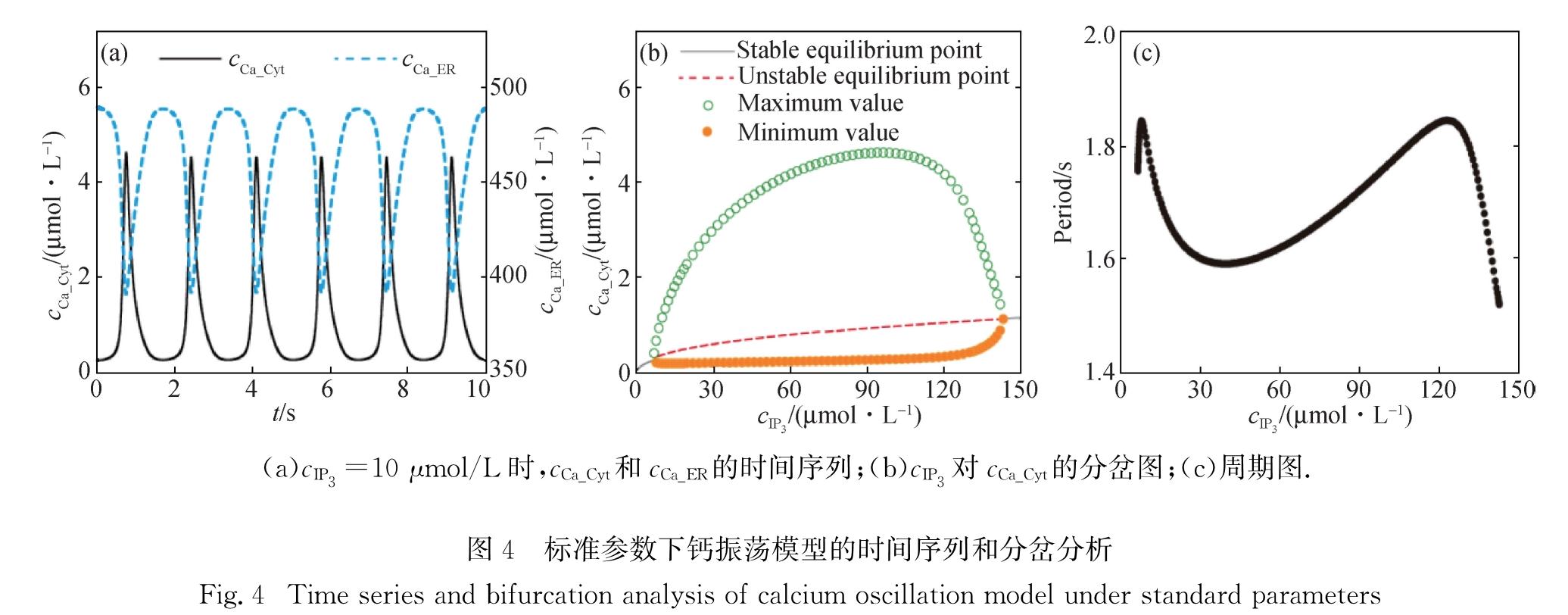
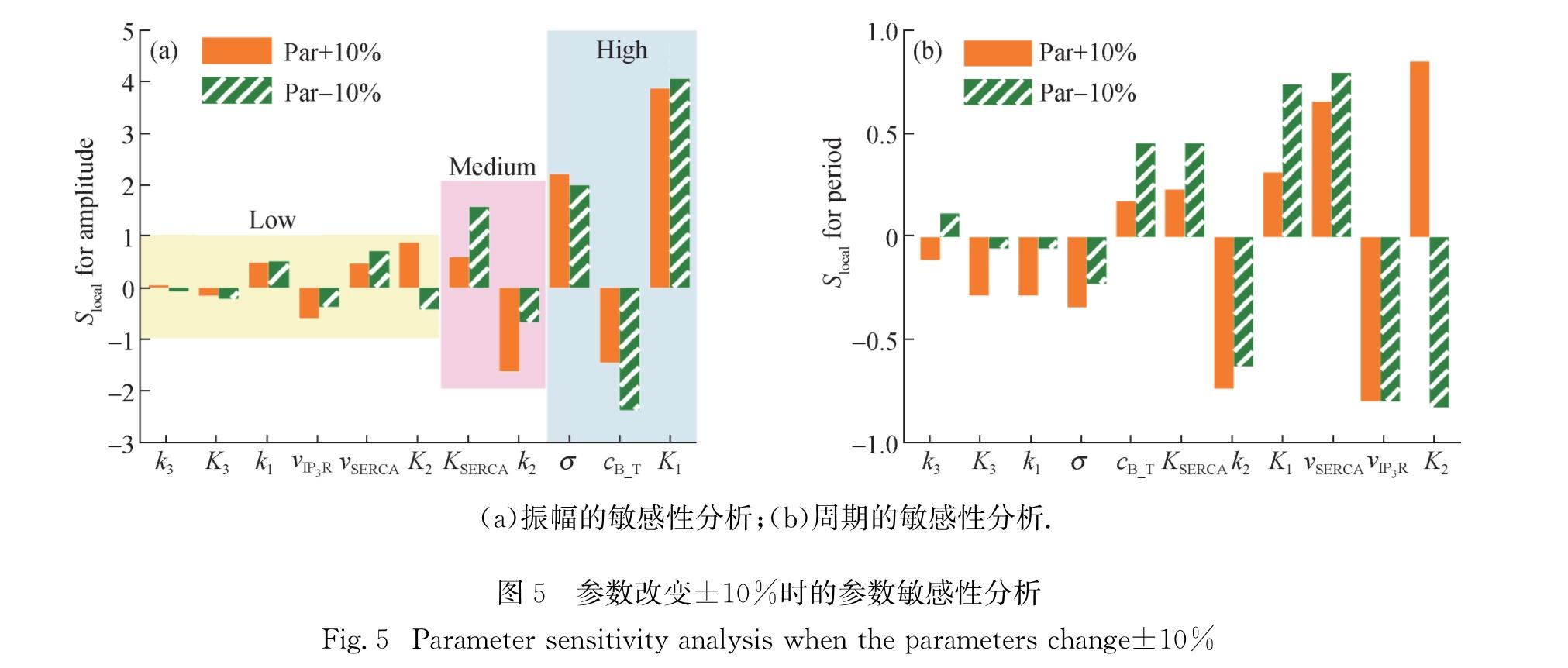
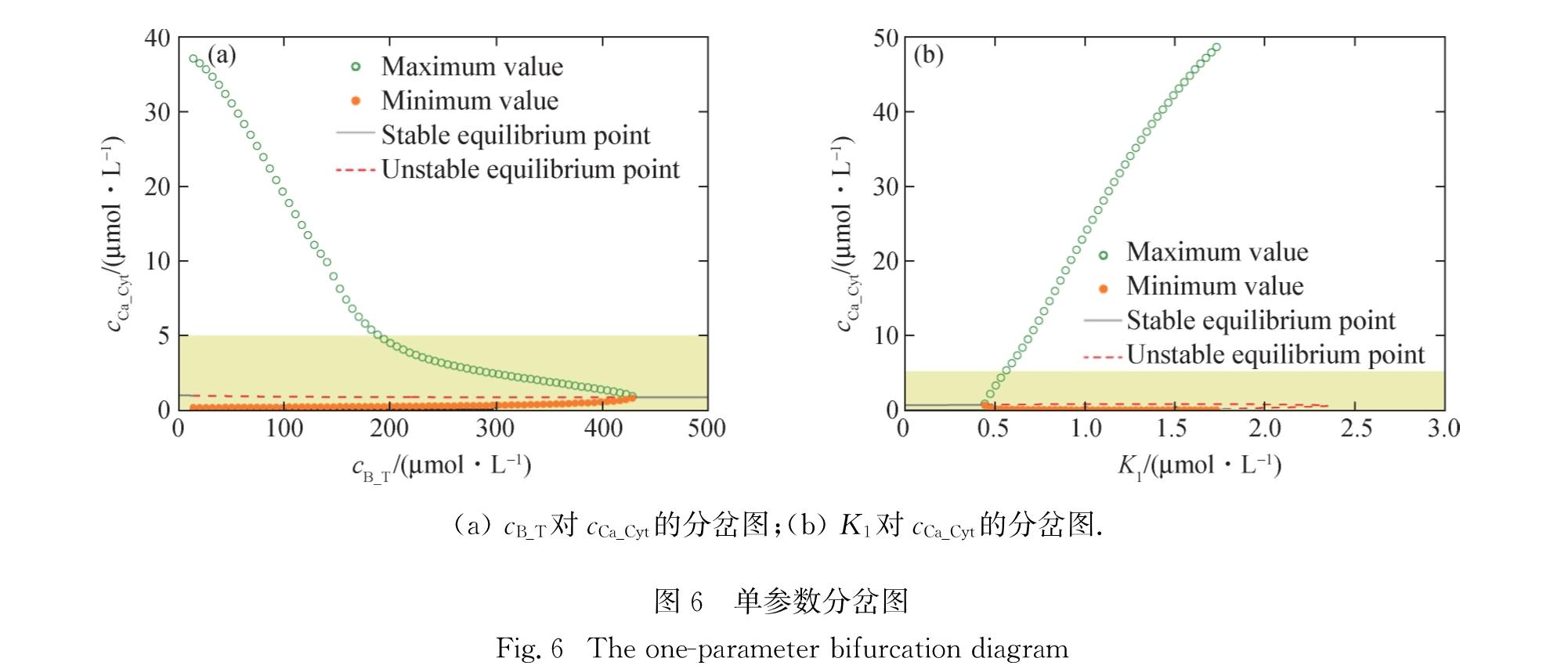
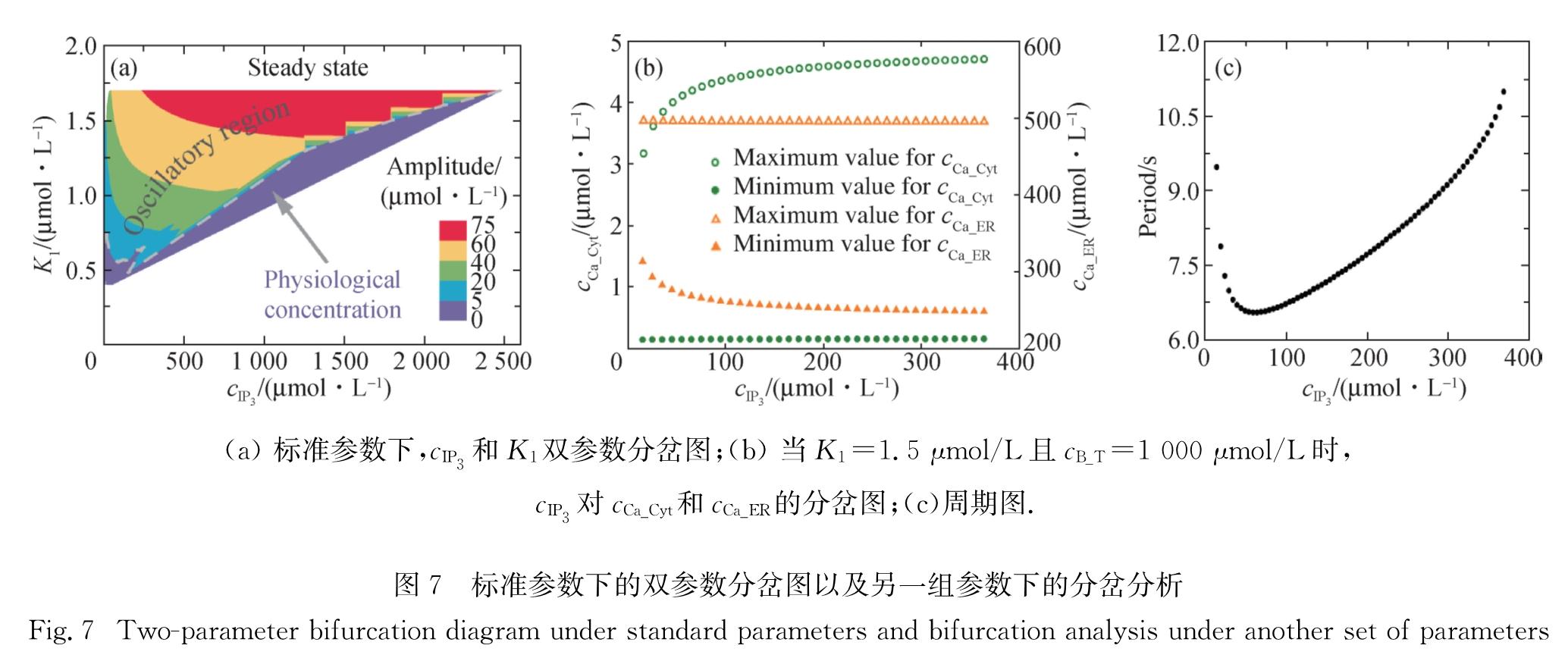
细胞质钙离子(Ca2+)常以浓度振荡变化的方式控制许多生理活动,内质网上的三磷酸肌醇受体(inositol 1,4,5-trisphosphate receptor,IP3R)通道是产生钙振荡的关键因素,因此研究其振荡机制具有重要意义.虽然钙振荡已被广泛建模,但大部分模型细胞质与内质网中钙振荡的范围与实验观测数据差异较大.通过整合最近的实验结果,基于IP3R通道活性对细胞质Ca2+浓度依赖的钟型曲线构建了一个新的钙振荡模型.模型结果除了可以很好地模拟实验中关于钙振荡范围的数据,还可重复多种实验现象.对模型进行参数敏感性分析后,进一步采用单参数分岔分析分别研究了两个高敏感参数,即细胞质内Ca2+缓冲蛋白的总浓度和IP3R通道激活Ca2+的解离常数(K1)对钙振荡的影响,发现它们对钙信号有相反的作用.对三磷酸肌醇(IP3)浓度和K1进行双参数分岔分析的结果表明K1对IP3能产生钙振荡的区域及其振幅有重要影响,揭示可以通过改变K1影响细胞对外界刺激的响应及其钙振荡模式.该研究有助于理解钙振荡机制,并可为后续理论研究提供框架.
Oscillations in cytosolic calcium concentration(cCa_Cyt)regulate a diverse range of cellular processes.Dynamic properties of inositol 1,4,5-trisphosphate receptor(IP3R)located on the endoplasmic reticulum are crucial for the generation of Ca2+ oscillations.Thus,it is of great interest to determine mechanisms governing such oscillations.Although Ca2+ oscillations have been modeled extensively,one major problem with almost all published models lies in that the oscillatory ranges of cCa_Cyt and calcium concentration in endoplasmic reticulum(cCa_ER)do not agree with recent experimental data.By integrating recent experimental results,a new calcium oscillation model is constructed based on bell-shaped dependence of IP3R gating on cCa_Cyt.The cCa_Cyt and cCa_ER obtained by the model lie in ranges of experimental results.In addition,the model can successfully reproduce several features described in experimental studies.After parameter sensitivity analysis of the model,the single-parameter bifurcation analysis is conducted to study the impact of two high-sensitivity parameters,i.e.,the total concentration of cytoplasmic Ca2+ binding protein and IP3R dissociation constant for activating Ca2+(K1)on Ca2+ oscillations. Results suggest that they exert an opposite effect on calcium signal.The result of two-parameter bifurcation analysis of IP3 concentration and K1 indicates that K1 plays a crucial role on the region of IP3 concentration that can generate Ca2+ oscillations and their amplitudes,revealing that K1 can affect the response of cells to external stimuli and the Ca2+ oscillatory pattern of the cells.This study not only helps to understand the mechanism of Ca2+ oscillations,but also provides a framework for subsequent theoretical research.