收稿日期:2018-12-05录用日期:2019-03-02
基金项目:国家自然科学基金(31360006,31760006); 西藏自治区自然科学基金(XZ 2018 ZR G-21)
通信作者:372807194@qq.com
基金项目:国家自然科学基金(31360006,31760006); 西藏自治区自然科学基金(XZ 2018 ZR G-21)
通信作者:372807194@qq.com
(1.西藏农牧学院植物科学学院,西藏 林芝 860000; 2.西藏高原生态研究所,西藏 林芝 860000)
(1.College of Plant Science,Tibet Agriculture & Animal Husbandry University,Linzhi 860000,China; 2.Research Institute of Tibet Plateau Ecology,Linzhi 860000,China)
DOI: 10.6043/j.issn.0438-0479.201812001
从西藏色季拉山不同海拔采集的323份标本中分离毛壳属(Chaetomium)及形态相似属真菌,共分离到29个菌株,通过形态学特征将其中的27个菌株分别鉴定为3属5种:球毛壳(C. globosum)、马德拉斯毛壳(C. madrasense)、近缘毛壳(C. subaffine)、绳生二叉毛壳(Dichotomopilus funicola)和粗壮围领毛壳(Colariella robusta).通过ITS rDNA和28S rDNA D1/D2区序列构建系统发育树,分离的29个菌株被划分为6个种.碱基变异分析结果表明,ITS rDNA序列较28S rDNA D1/D2区序列变异更大,结合这两段基因序列可进行毛壳属及形态相似属真菌属内种间的系统发育研究.这些结果可丰富西藏毛壳属及形态相似属真菌资源库,为开发毛壳属及形态相似属真菌代谢产物奠定基础.
323 samples were collected from different altitudes of Sejila Mountains in Tibet,and 29 strains of Chaetomium and Chaetomium-like fungi were isolated.The results showed that 27 strains of them were identified as 3 genera 5 species,C.globosum,C.madrasense,C.subaffine,Dichotomopilus funicola and Colariella robusta.Phylogenetic tree was constructed based on sequences of ITS rDNA and 28S rDNA D1/D2 region,showing that the 29 isolated strains were divided into 6 groups.The results of base mutation analysis showed that the ITS rDNA region had more variations than that of the 28S rDNA D1/D2 region,and the combination with these two gene sequences could be used in the phylogenetic study between species within genus of Chaetomium and Chaetomium-like fungi.Taken together,this study enriched the resources of Chaetomium and Chaetomium-like fungi in Tibet and laid a foundation for exploring metabolites of Chaetomium and Chaetomium-like fungi.
毛壳属(Chaetomium Kunze)真菌(简称毛壳菌)隶属于子囊菌门(Ascomycota)核菌纲(Pyrenomycetes)粪壳菌目(Sordariales)毛壳菌科(Chaetomiaceae)[1],是毛壳菌科最大的属,据《真菌字典》(第10版)记载共81个种[2].我国对毛壳菌的分类研究始于20世纪50年代末,陈庆涛[3]报道了2个新种,戴芳澜[4]记录了该属30个种(含异名),孙广宇等[5]报道了3个新种和4个新记录种,谭悠久[6]发现了1个新种和10个新记录种,王雪薇[7]报道了31个种,其中包括2个新种和5个我国新记录种,刘富江[8]报道了14个种,其中1个为我国新记录种.目前,我国发现并记录的毛壳菌共有31种.
毛壳菌广泛分布于自然界含纤维素的各种基质上,可有效降解纤维素和木质素等难降解的大分子有机物,并在生物防治中发挥重要作用.现已发现球毛壳(C. globosum)和角毛壳(C. cupreum)可以用来防治多种植物病害[9-10]; 球毛壳菌菌肥可提高土壤酶活性[11],促进小麦根系发育和提高其抗旱性[12],提高杨树叶片的光合效率[13]; 此外,毛壳菌的代谢产物还可诱导植物增强抗病能力,提高作物产量[14].
20世纪60年代以后,分子生物学方法被广泛应用于真菌分类和鉴定,其中ITS rDNA序列被推荐为真菌的通式性DNA条形码[15-16].该条形码同样适用于毛壳菌的分类鉴定,2016年Wang等[17]结合多基因系统发育分析,重新确定了毛壳菌中36个种的分类地位,将这36个种分别划分到13个属内(作为其形态相似属),这是毛壳菌分类史上的一次重大变化.
青藏高原被誉为“世界第三极”,地形复杂,土壤种类繁多,植被丰富,特别适合土壤真菌的生长和繁殖,真菌多样性指数和丰富度较高,长期以来一直受到中外真菌学家们的关注.徐阿生等[18]和张惠等[19]对西藏大型真菌进行了调查,蔡晓布等[20]对西藏丛枝菌根真菌的多样性进行了研究.目前已经记录的西藏真菌总计185科551属2 599种,占我国真菌总数的18.5%[21],占世界真菌总数的2.4%[22],这些数据说明西藏是重要的菌种资源库.色季拉山位于我国藏东南雅鲁藏布江大峡谷西北侧,主峰海拔高度为5 200 m,属于藏东南半湿润区与湿润区的过渡地带,其动植物和菌物资源很丰富,是西藏生物多样性最丰富的山体之一.近年来,我国学者也开展了一些关于西藏色季拉山真菌资源的研究,如:刘小娇等[23]对色季拉山区红菇科大型真菌进行了调查和采集,鉴定出红菇属(Russula)15种、乳菇属(Lactarius)8种; 邓丽君等[24]对色季拉山丝膜菌属(Cortinarius)真菌进行了调查和采集,鉴定出丝膜菌属真菌16种.
目前,关于西藏毛壳属及形态相似属真菌的研究报道还较少.刘述春等[25]对来源于西藏林芝地区的一株毛壳菌的固体发酵提取物进行了生物活性筛选,发现了新的活性化合物新聚酮类; Guo等[26]从西藏林芝地区油松和巨柏根际土壤中分离得到3株毛壳菌并发现1个新种.上述研究表明特殊生境是微生物新种和活性天然产物的重要来源.本研究从色季拉山不同海拔采集植物残体、土壤和动物粪便等标本,进行毛壳属及形态相似属真菌的分离和鉴定,分析色季拉山毛壳属及形态相似属真菌资源现状,以期为筛选西藏无公害生产的毛壳属及形态相似属真菌的菌株奠定基础.
323份样品采集于西藏色季拉山沿线2 000~3 792 m 海拔高度的土壤、植物残体和动物粪便等.
1.1.2 供试培养基[6]
分离培养基为马丁氏培养基,鉴定培养基为玉米粉培养基(CMA).
1.2.1 菌株分离与纯化[6]
树枝、树叶、朽木、杂草和动物粪便样品采用组织分离法:将实验材料切成4~5 mm的组织小块,放入60%(体积分数)的乙醇中消毒6~8 min,无菌条件下用无菌水连续漂洗3次; 将组织小块移至马丁氏培养基平板上,每皿内4~5块,25 ℃恒温培养,3~4 d后观察结果; 解剖镜下挑取单个子囊果接种于CMA平板上,进行菌株的纯化和观察.土壤样品采用稀释分离法.
形态鉴定参考Arx系统[27]和Wang等[17]的研究,将菌株接种于CMA平板上,28 ℃黑暗培养,定期观察直至子囊果完全成熟.采用莱卡(Leica)显微镜DM5000观察,每个菌株研究和测量20个个体.
DNA的提取主要参考Nakada等[28]的方法,略有改进.ITS rDNA序列的PCR引物对为ITS1-F(5'-CTTGGTCATTTAGAGGAAGTAA-3')和ITS4(5'-TCCTCCGCTTATTGATATGC-3'),28S rDNA D1/D2区序列的PCR引物对为NL1(5'-GCATATCAAT
AAGCGGAGGAAAAG-3')和NL4(5'-GGTCCGTGT
TTCAAGACGG-3'),由北京六合华大基因科技有限公司武汉分公司合成.PCR产物由上海英骏生物技术有限公司用ABI 3730 DNA测序仪直接进行序列测定.
用SequenceMatrix(Windows-1.7.8)软件进行菌株ITS rDNA和28S rDNA D1/D2区序列的拼接.利用DnaSP 5.0软件对序列进行DNA多态性分析,分别统计变异位点数(S)、单倍型数目(H)、单倍型多样度(Hd)、核苷酸多样度(Pi)和核苷酸平均差异数(k).将分离所得29个菌株的ITS rDNA和28S rDNA D1/D2区序列的测序结果提交至GenBank并申请接受号; 再利用BLAST(https:∥blast.ncbi.nlm.nih.gov)进行同源序列查找,选择所需相关种的序列,并以小囊菌(Microascus trignosporus)为外群,用MEGA 6.0软件构建系统进化树.
从西藏色季拉山沿线2 000~3 792 m海拔高度采集的323份土壤、植物残体、动物粪便中,共分离到29株毛壳属及形态相似属真菌,分离率为8.98%.将分离到的29个菌株的ITS rDNA和28S rDNA D1/D2区序列的测序结果提交至GenBank并申请接受号,如表1所示.
分离到的29株毛壳属及形态相似属真菌中27株被鉴定为5个种:17株为球毛壳(C. globosum),菌株号为20-3-1、22-1、22-3-1、22-3-2、22-10、23-41、24-3-1、24-3-2、Z24-5、24-9、24-15-1、24-15-2、Z24-23-1、25-9-1、25-9-2、Z27-11-1和Z27-11-2,占分离所得毛壳菌菌株总数的58.62%,为优势种群; 7株为绳生二叉毛壳(Dichotomopilus funicola),菌株号为2-2-1、2-2-2、Z18-1、22-3-3、Z24-2、24-28和32-5,占分离所得毛壳菌菌株总数的24.14%; 马德拉斯毛壳(C. madrasense)1株,菌株号为Z24-13-1; 近缘毛壳(C. subaffine)1株,菌株号为Z24-12; 粗壮围领毛壳(Colariella robusta)1株,菌株号为LZT0008.5个种的代表菌株具体形态特征如图1~5所示.还有2个菌株Z24-11和Z30-4通过形态学特征尚不能准确鉴定到种.
C. globosum Kunze-Mykol. Hefte 1:16.1817(Fig.1)
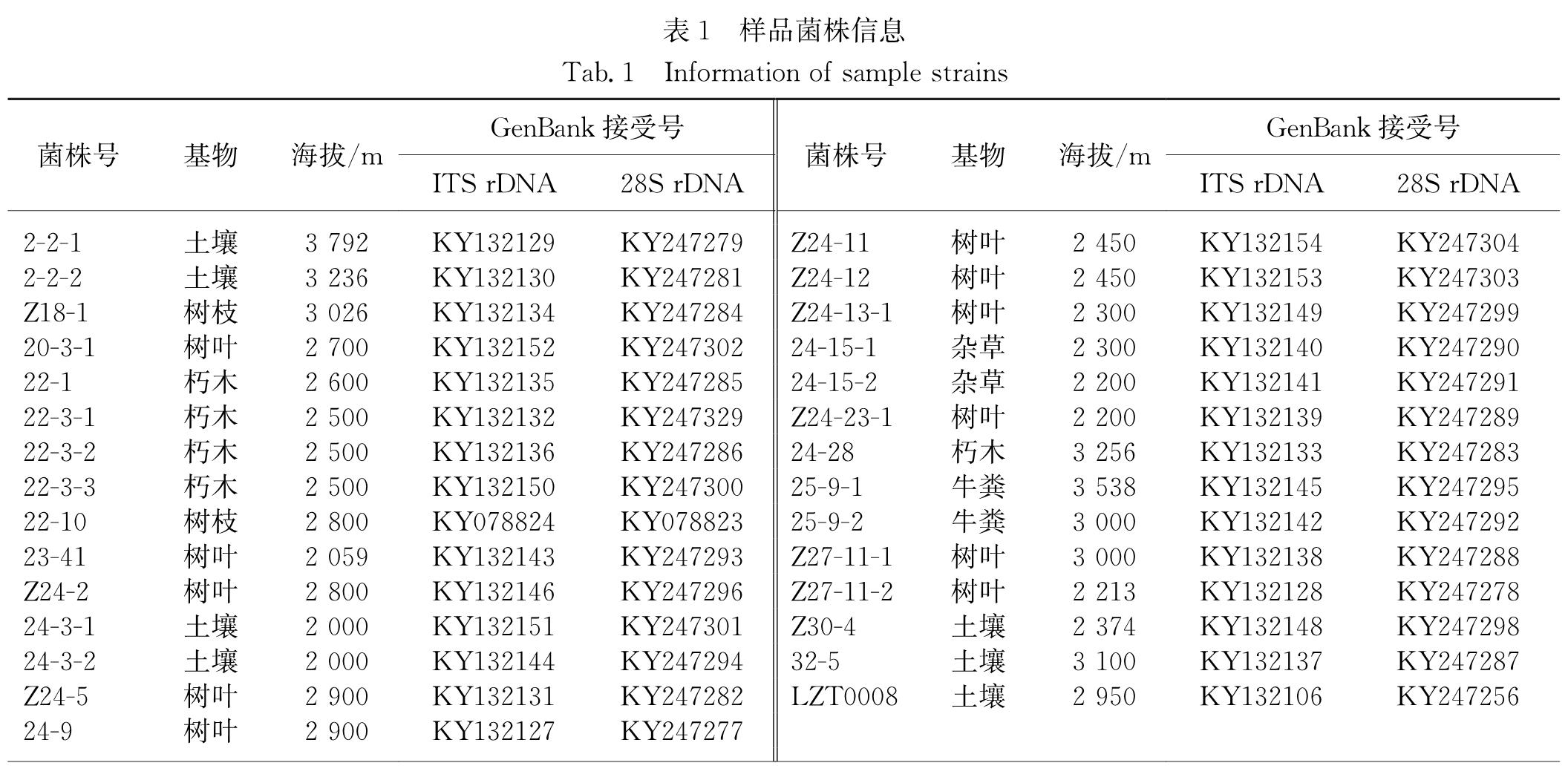
表1 样品菌株信息
Tab.1 Information of sample strains
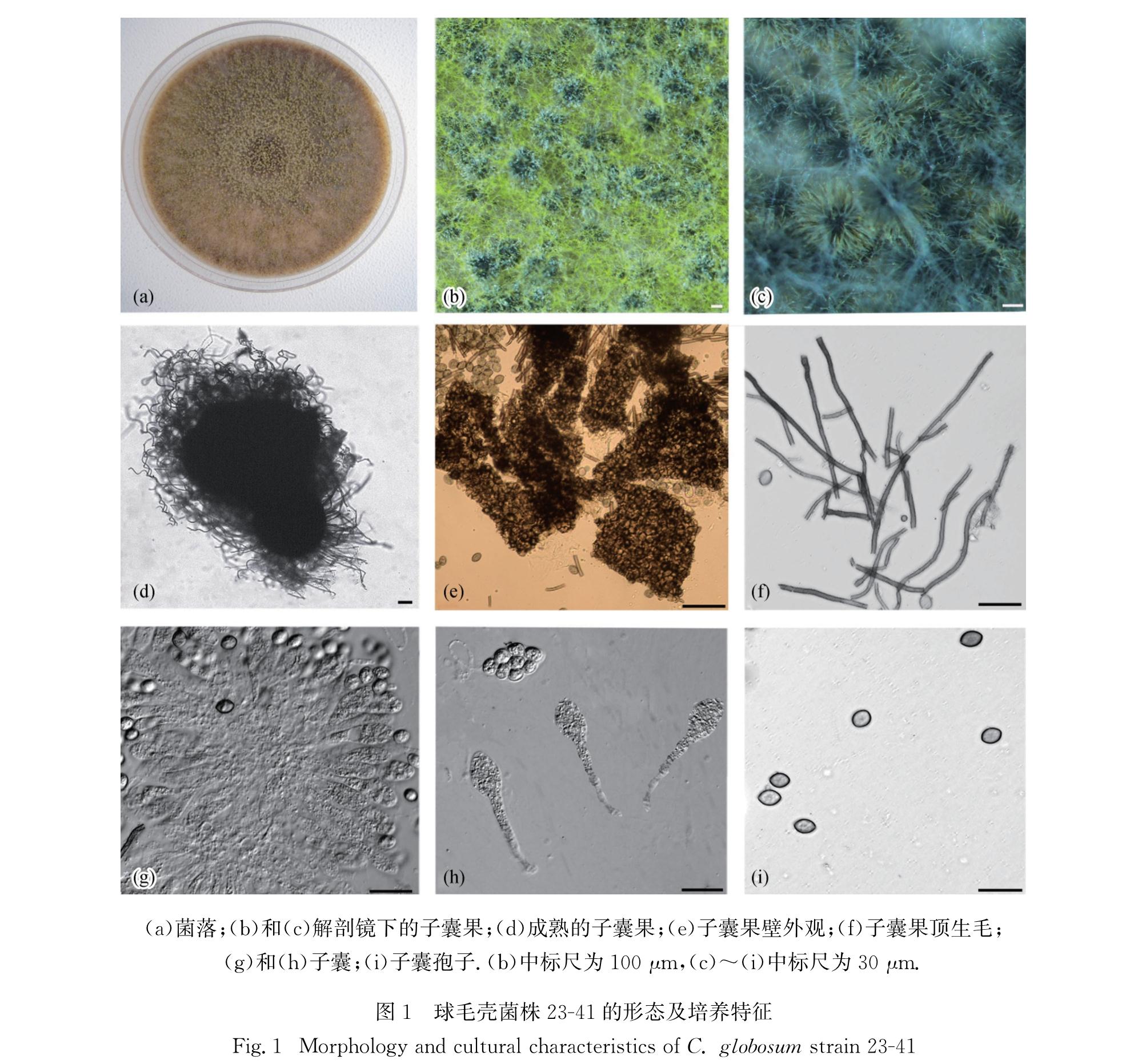
图1 球毛壳菌株23-41的形态及培养特征
Fig.1 Morphology and cultural characteristics of C. globosum strain 23-41
菌落在CMA平板上的直径日平均生长量为7~8 mm,气生菌丝呈浅黄绿色至橄榄色,产生黄色至黄绿色分泌物; 子囊果不埋生,球形至椭圆形,直径170~290 μm,有固定孔口,7~9 d开始成熟,反射光下呈灰褐色至浅橄榄色; 果壁黄褐色,果壁细胞呈暗褐色交织状; 子囊果顶生附属丝密生,褐色,直或略带弯曲,具疣,具隔,基部宽度3.0~4.5 μm,最长可达500 μm; 子囊簇生,棍棒状,具柄,含8个子囊孢子,易消解,含柄部分长轴50~80 μm,短轴10~16 μm; 子囊孢子成熟时呈浅褐色,内含多个油滴,柠檬形,两侧平滑,两头稍尖具突起,长轴10~12 μm,短轴6~8 μm,有单个顶生萌发孔[29].
C.madrasense Natarajan-Proc.Indian Acad.Sci., B, 74:255.1971(Fig.2)
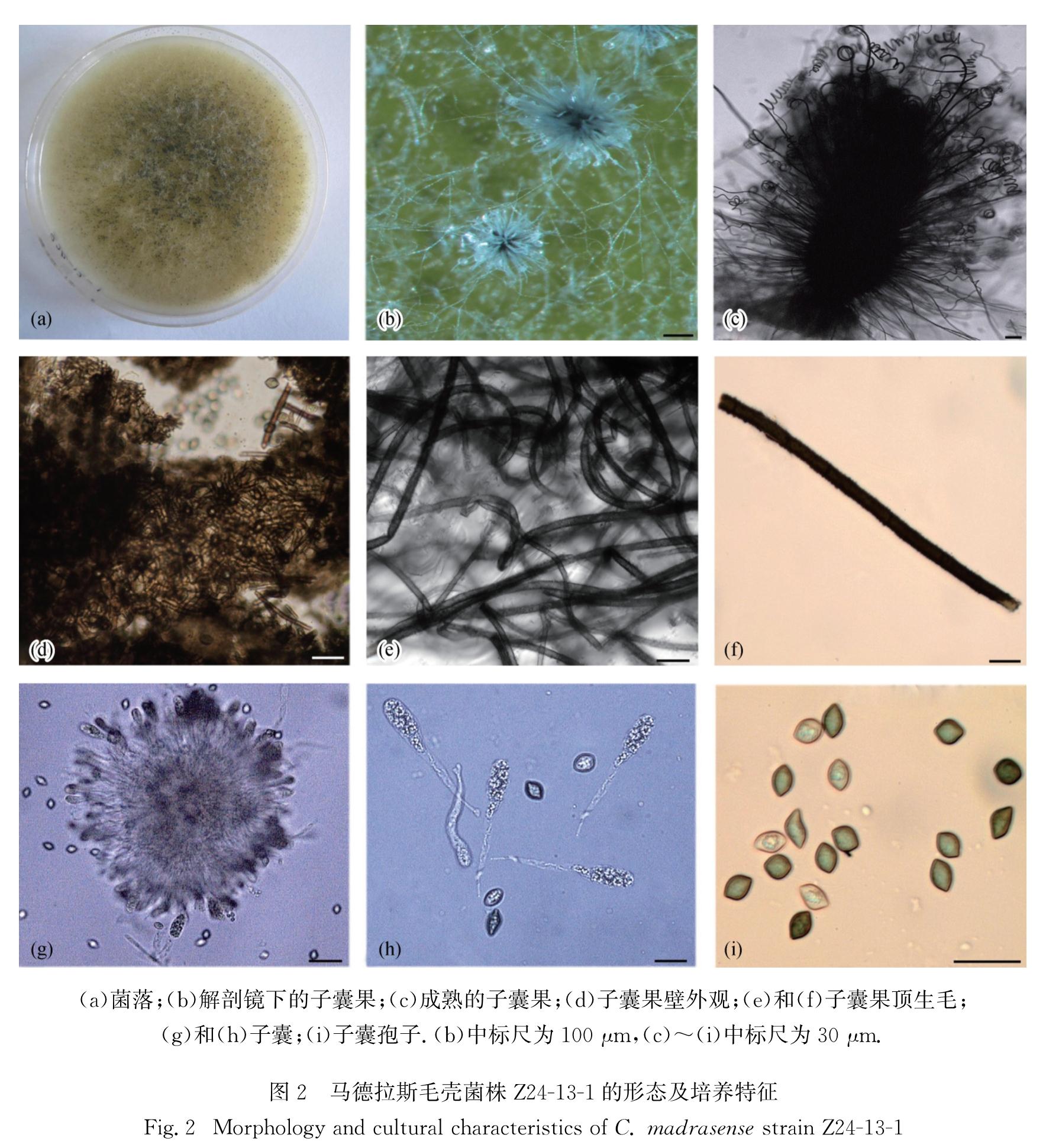
图2 马德拉斯毛壳菌株Z24-13-1的形态及培养特征
Fig.2 Morphology and cultural characteristics of C. madrasense strain Z24-13-1
菌落在CMA平板上生长稀疏,呈浅黄绿色,无明显分泌物产生; 子囊果不埋生,反射光下呈橄榄绿色至黑褐色,卵圆形,具固定孔口,直径200~310 μm,子囊果果壁呈褐色交织状; 子囊果顶生附属丝弯曲,小波浪状或顶端略呈螺旋形,浅褐色,分隔不明显,具疣,基部宽度2~4 μm; 子囊簇生,棍棒状,具柄,内含8个子囊孢子,易消解,无柄部分长轴24.6~29.7 μm,短轴10.9~16.0 μm; 子囊孢子呈柠檬形,成熟时褐色,两侧略扁不对称,一侧常有一个明显凸起,内含多个油滴,长轴10.3~13.5 μm,短轴6.6~9.7 μm,有单个顶生萌发孔.
C.subaffine Sergejeva-Not.Syst.Sect.Crypt.Inst.Bot.Acad.Sci.U.S.S.R.14:148.1961(Fig.3)
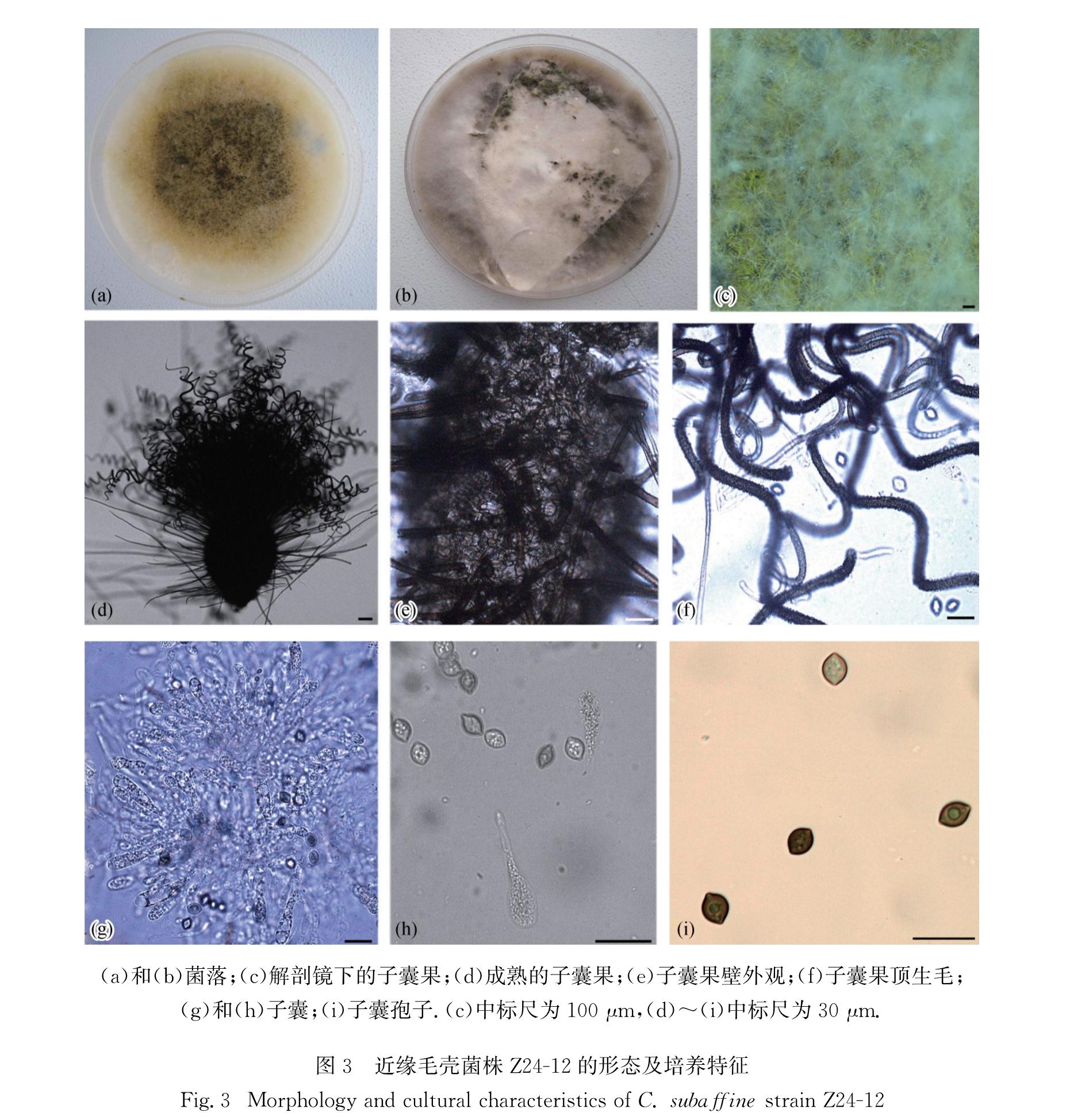
图3 近缘毛壳菌株Z24-12的形态及培养特征
Fig.3 Morphology and cultural characteristics of C. subaffine strain Z24-12
菌落在CMA平板上呈白色至浅褐色,略有黄色分泌物; 子囊果不埋生,卵圆形至安培瓶形,220~380 μm,有固定孔口,成熟较慢,培养至12 d后才开始成熟,但在滤纸培养基上产果较容易,气生菌丝丰富,反射光下呈深橄榄色至浅褐色,果壁呈深褐色交织状; 子囊果顶生附属丝密生,褐色,具疣,有分隔,螺旋状弯曲,基部宽度4~5 μm; 子囊丛生,棍棒状,易消解,具柄,含8个子囊孢子,无柄部分长轴41~68 μm,短轴10~16 μm; 子囊孢子呈褐色,柠檬形或宽纺锤形,两侧平滑,两头略尖,长轴10~13 μm,短轴8~10 μm,有单个明显的顶生萌发孔.
D. funicola,异名:粪生毛壳C. funicola Cooke-Grevillea 1:176.1873(Fig.4)
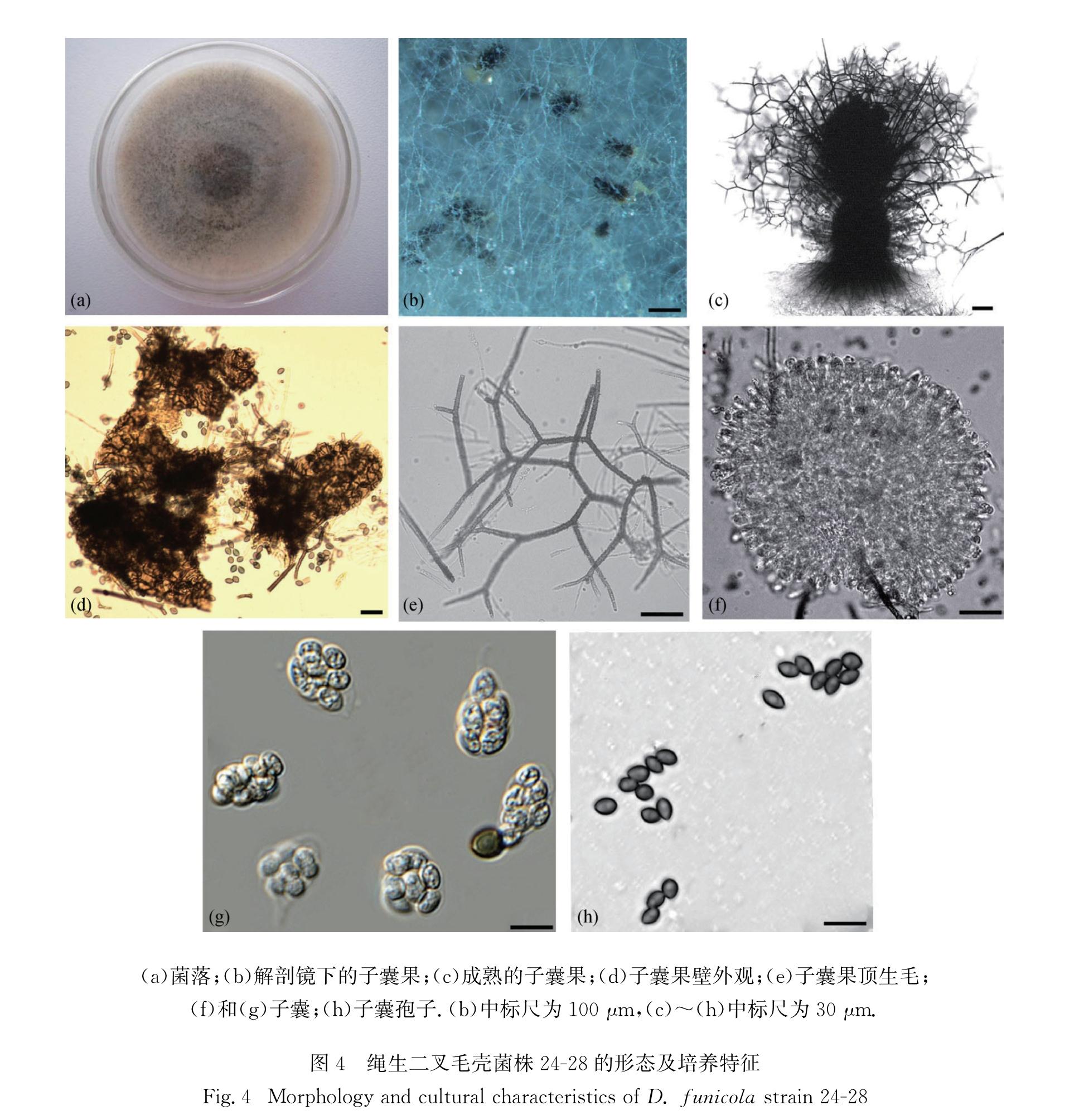
图4 绳生二叉毛壳菌株24-28的形态及培养特征
Fig.4 Morphology and cultural characteristics of D. funicola strain 24-28
菌落在CMA平板上气生菌丝生长稀疏,呈白色至浅黄色,略产生浅茶色分泌物; 子囊果不埋生,球形至卵圆形,直径176~234 μm,有固定孔口,10 d后开始成熟,反射光下呈暗褐色至黑色; 子囊果附属丝聚集顶生,多轮二叉状分枝,附属丝反射光下呈暗褐色,具疣,具隔,基部宽度5~6 μm; 子囊簇生,棍棒状,极易消解,具长柄,内含8个子囊孢子,无柄部分长轴20~32 μm,短轴8~12 μm; 子囊孢子呈褐色,卵圆形,两侧平滑对称,一头稍尖,长轴4.0~5.5 μm,短轴5.7~7.2 μm,有单个顶生萌发孔.
C. robusta,异名:粗壮毛壳C. robusum Ames-Monograph Chaetomiaceae p.35.1963(Fig.5)
菌落在CMA平板上不产生气生菌丝; 子囊果稍埋生,坛形至长椭圆形,有固定孔口,直径 180~280 μm,最高可达600 μm,7~10 d后开始成熟,反射光下呈金属灰色至黑褐色; 子囊果果壁细胞成熟后呈深褐色,不规则形,大小7.9~10.6 μm,果壁细胞排列似花瓣状,靠近孔口的果壁细胞呈细长栅栏形,整齐排列成3~7层,厚度约57 μm; 子囊果顶生附属丝褐色,具隔,具疣,下部刚直,顶端呈波浪状至螺旋状; 子囊簇生,棍棒状,易消解,柄较长,内含8个子囊孢子,无柄部分长轴30~40 μm,短轴10~16 μm; 子囊孢子呈柠檬形,成熟后浅褐色,两侧平滑对称,两头稍尖具突起,长轴5.5~7.5 μm,短轴4.0~6.0 μm,有单个顶生萌发孔.
分别扩增29株毛壳菌的ITS rDNA和28S rDNA D1/D2区2个基因片段,ITS rDNA扩增获得538 bp,28S rDNA D1/D2区扩增获得511 bp,拼接序列总长为1 049 bp,共有63个变异位点,11种单倍型,单倍型多样度为0.845,核苷酸多样度为0.020 22,核苷酸平均差异数为19.185,ITS rDNA序列的多样性指标值均大于28S rDNA D1/D2区(表2).
从GenBank中选择18个相关模式种的ITS rDNA和28S rDNA D1/D2区序列(表3),与分离的
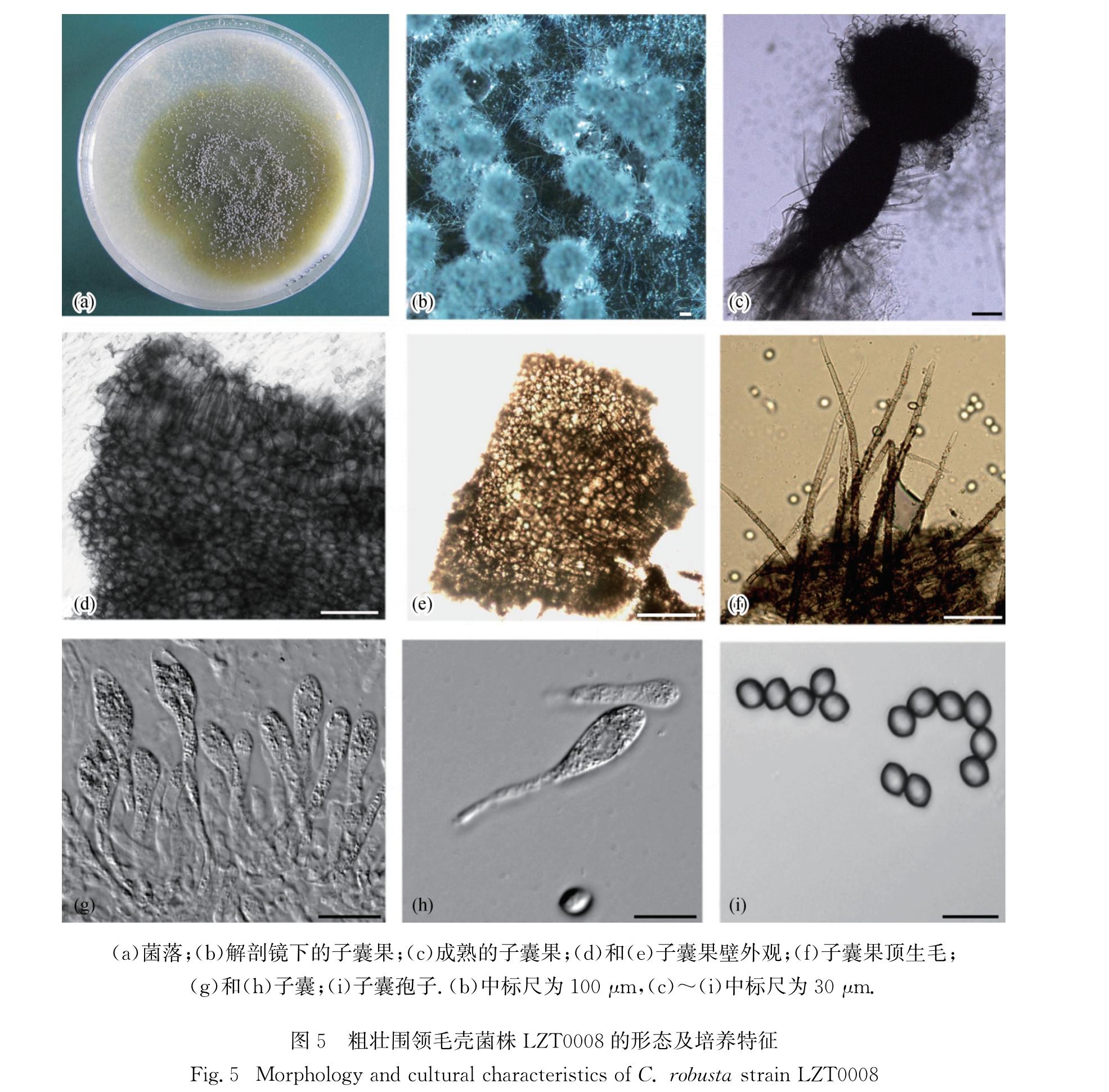
图5 粗壮围领毛壳菌株LZT0008的形态及培养特征
Fig.5 Morphology and cultural characteristics of C. robusta strain LZT0008
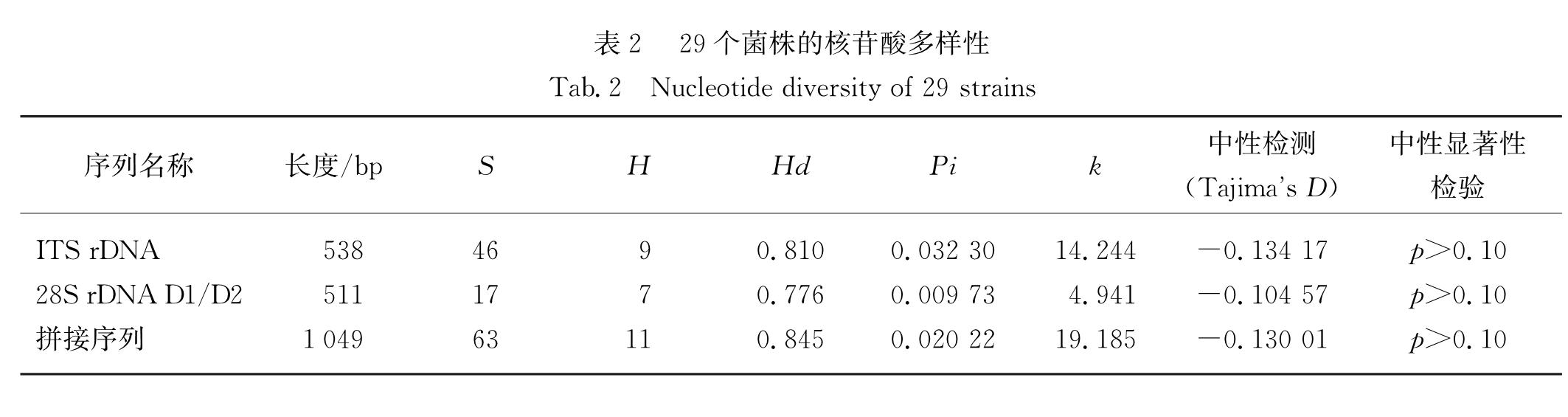
表2 29个菌株的核苷酸多样性
Tab.2 Nucleotide diversity of 29 strains
29个菌株序列一起构建色季拉山毛壳属及形态相似属真菌的系统发育树(图6).
结果显示分离的29个菌株被划分在6个分支中:第1分支包括了形态学上鉴定为球毛壳的12个菌株(22-1、22-3-2、22-10、23-41、24-3-1、24-3-2、24-15-2、25-9-1、25-9-2、Z27-11-2、22-3-1、Z24-5)和4个球毛壳模式菌株(CGMCC 3.12922、CGMCC 3.9994、CBS 105.40、CBS 148.51),自举值达97%; 第2分支包括了形态学上鉴定为马德拉斯毛壳的1个菌株(Z24-13-1)和2个马德拉斯毛壳模式菌株(CBS 315.74和CBS 126663),自举值达86%; 第3分支包括了形态学上鉴定为球毛壳的5个菌株(24-15-1、24-9、20-3-1、Z24-23-1、
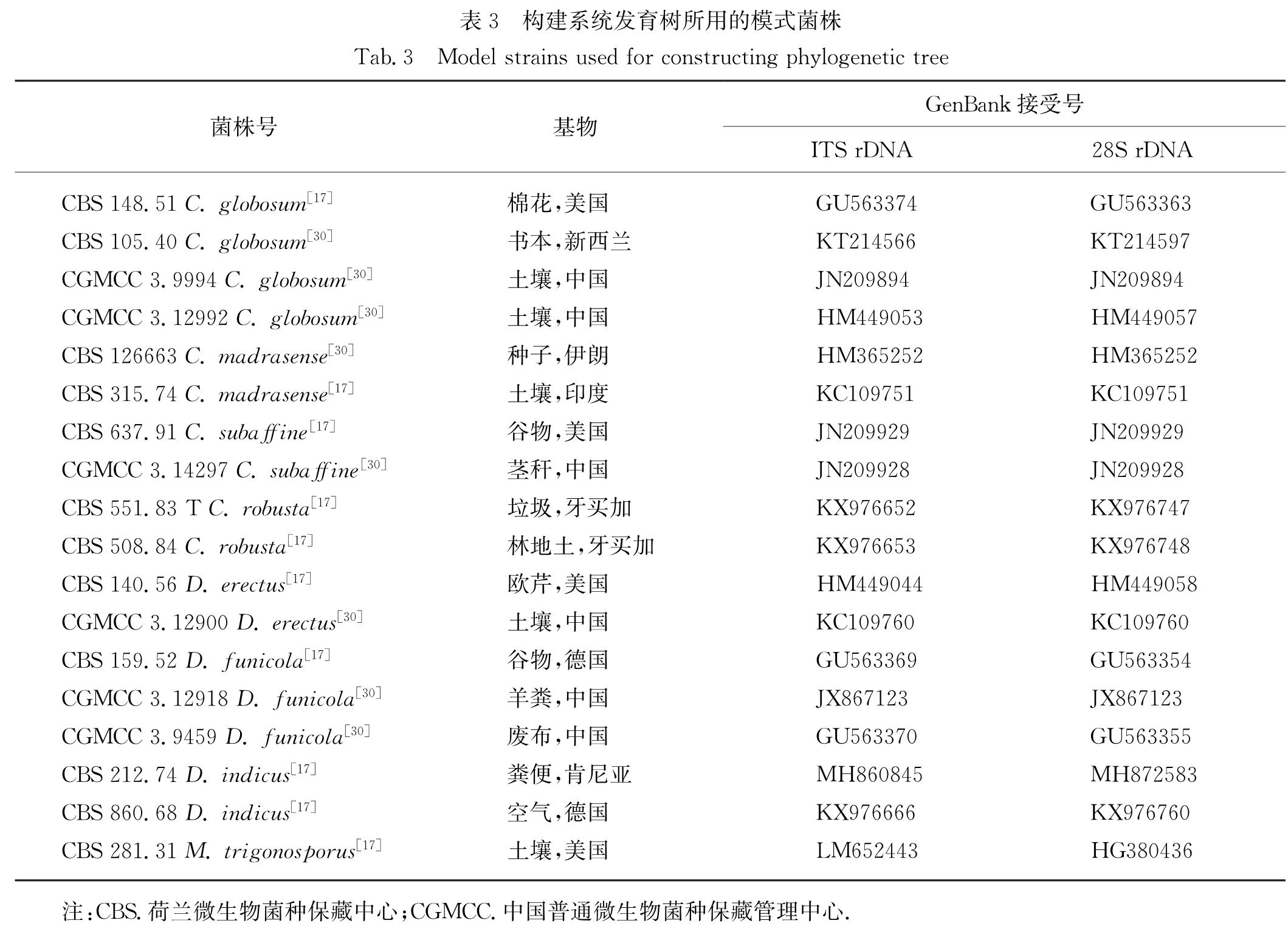
表3 构建系统发育树所用的模式菌株
Tab.3 Model strains used for constructing phylogenetic tree
注:CBS.荷兰微生物菌种保藏中心; CGMCC.中国普通微生物菌种保藏管理中心.
Z27-11-1)、形态学上鉴定为近缘毛壳的1个菌株(Z24-12)和2个近缘毛壳模式菌株(CBS 637.91和CGMCC 3.14297),还包括形态学上不能准确鉴定到种的1个菌株(Z24-11),自举值达88%; 第4分支包括了形态学上不能准确鉴定到种的1个菌株(Z30-4)和2个直立二叉毛壳模式菌株(CGMCC 3.12900和CBS140.56),自举值达99%; 第5分支包括了形态学上鉴定为绳生二叉毛壳的7个菌株(2-2-1、2-2-2、22-3-3、24-28、32-5、Z18-1、Z24-2)和3个绳生二叉毛壳模式菌株(CBS159.52、CGMCC 3.12918、CGMCC 3.9459),自举值达99%; 第6分支包括了形态学上鉴定为粗壮围领毛壳的1个菌株(LZT0008)和2个粗壮围领毛壳模式菌株(CBS 508.834和CBS 551.83 T),自举值达99%.
从系统发育树可以看出,ITS rDNA和28S rDNA D1/D2区序列可以用来区分毛壳属及形态相似属真菌中的一些形态差异较大的种,如球毛壳、绳生二叉毛壳、粗壮围领毛壳、马德拉斯毛壳被划分在不同的分支上; 但对于形态学上极为相似的种,如球毛壳和近缘毛壳不能很好地区分,这2个种的部分菌株也被聚在一个分支上.
综上,形态学上不能准确鉴定的2个菌株Z24-11和Z30-4,通过分子生物学数据分别被鉴定为近缘毛壳和直立二叉毛壳; 通过形态特征鉴定为5个种的27个菌株,结合ITS rDNA和28S rDNA D1/D2区序列也被鉴定为5个种,即球毛壳、马德拉斯毛壳、近缘毛壳、粗壮围领毛壳、绳生二叉毛壳,可见形态鉴定结果与分子鉴定结果保持高度一致.
本研究从西藏色季拉山采集土壤、植物残体和动物粪便样品323份,共分离到毛壳属及形态相似属真菌29株,分离率仅为8.98%,该分离率与很多学者的研究结果相近,如:王雪薇[7]从我国17个省(市、自治区)采集标本1738份,获得136株菌,分离率为7.82%; 刘富江[8]从我国11个省(市、自治区)采集标本1098份,获得63株菌,分离率为5.74%.从本研究的种群丰富度来看,球毛壳和绳生二叉毛壳为优势种群.
分支点数值表示自举值,标尺表示每个核苷酸位点上的0.01替换值.
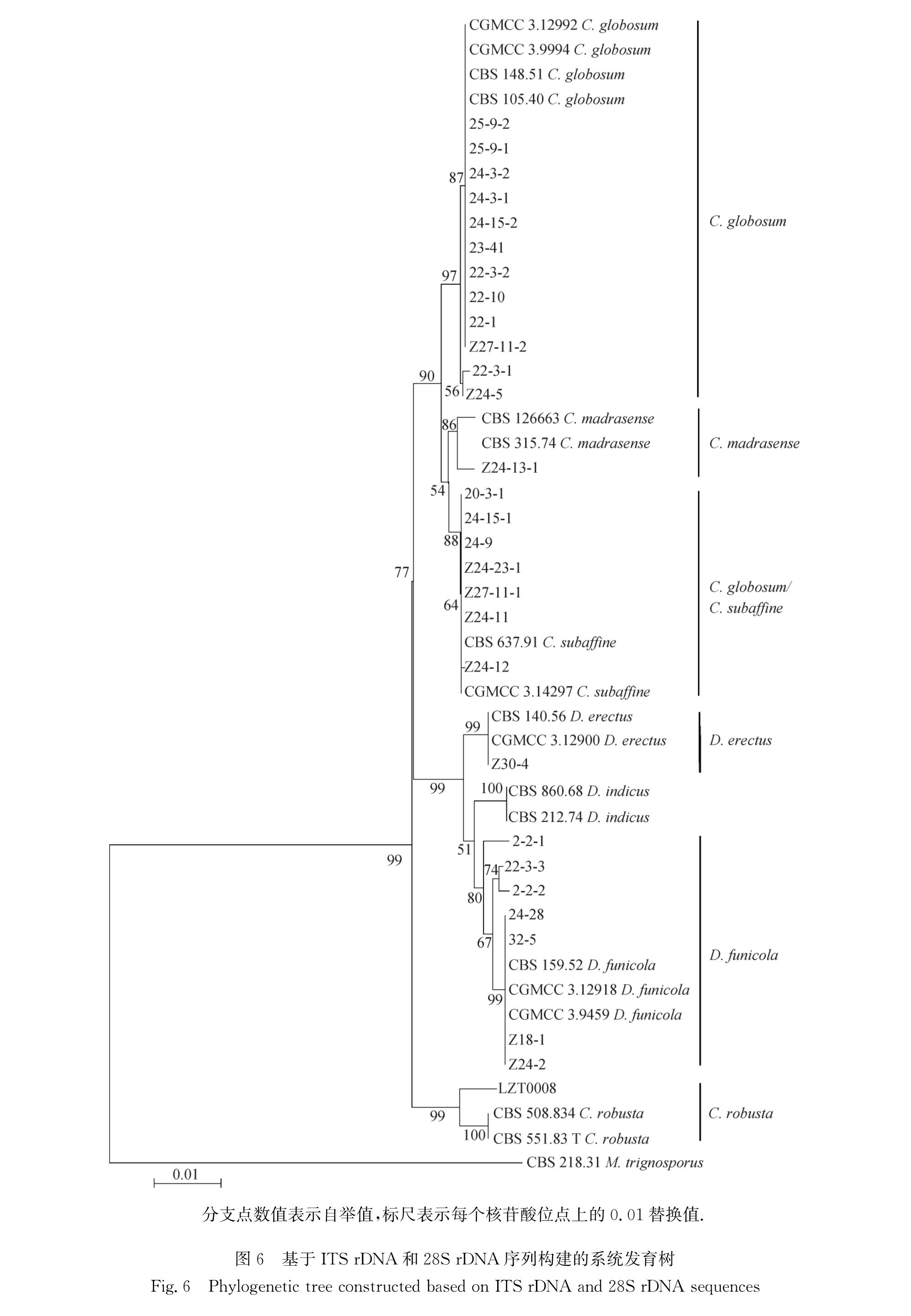
图6 基于ITS rDNA和28S rDNA序列构建的系统发育树
Fig.6 Phylogenetic tree constructed based on ITS rDNA and 28S rDNA sequences
从本研究中碱基变异的结果分析,ITS rDNA序列总体在碱基变异位点、单倍型、单倍型多样度、核苷酸多样度和核苷酸平均差异数等方面较28S rDNA D1/D2区变异更大,适合种级以上的分析; 而已有研究显示5.8S rDNA及其两端的ITS序列不加入成熟核糖体,所受到的选择压力较小,进化速率较快,可应用于真菌中属内种间的系统发育研究,作为真菌分子鉴定和多样性研究的DNA条形码[31-32].
在本研究中构建的系统发育树上,球毛壳的17个菌株分别被划分在2个分支上,说明球毛壳菌的不同菌株之间存在变异,这与Arx等[26]和王雪薇[7]的研究结果一致,认为球毛壳是一个变异较大的类群,几乎每个菌株的菌丝渗出物颜色、子囊果顶生附属丝形态、子囊孢子大小等方面都不完全相同.绳生二叉毛壳与印度二叉毛壳(D. indicus)在系统发育树上明显被划分在不同的分支上,这与王雪薇[7]的研究结果一致,两者的子囊果、子囊孢子的形态和大小等方面非常相似,其形态的主要区别在于子囊果顶生附属丝,说明ITS rDNA和28S rDNA D1/D2区序列可以用于区分形态上难以区分的绳生二叉毛壳与印度二叉毛壳.第3分支上将部分球毛壳菌株和近缘毛壳聚为一支,这与刘富江[8]的研究结果一致,近缘毛壳和球毛壳形态特征极为相似,只是前者的子囊果和子囊孢子较大,后者的子囊孢子一般小于12 μm,说明仅用ITS rDNA和28S rDNA D1/D2区序列还不能明显区分这两个形态上相似的种,需要参考更多的基因序列进行区分.本研究中2个通过形态学特征不能准确鉴定的菌株,通过ITS rDNA和28S rDNA D1/D2区序列可以进行初步分子鉴定,说明结合多基因片段有助于提高菌株鉴定的准确度.
综上,本研究从西藏色季拉山采集的323份样品中共分离到29株毛壳属及形态相似属真菌,其中球毛壳和绳生二叉毛壳为优势种群.通过形态学特征将29个菌株鉴定为5个种和2个未知种,再通过ITS和28S rDNA序列将29个菌株划分为6个种,为丰富西藏毛壳菌资源库和开发新的毛壳菌菌肥和真菌代谢产物奠定了基础.
