收稿日期:2018-06-14 录用日期:2018-08-04
< br/>基金项目:国家重点研发计划(2018YFC1406301); 国家自然科学基金(41476114); 厦门市海洋经济发展专项(14CZY042HJ16); 自然资源部“全球变化与海气相互作用”专项
< br/>通信作者:guodh@xmu.edu.cn < br/>
< br/>通信作者:guodh@xmu.edu.cn < br/>
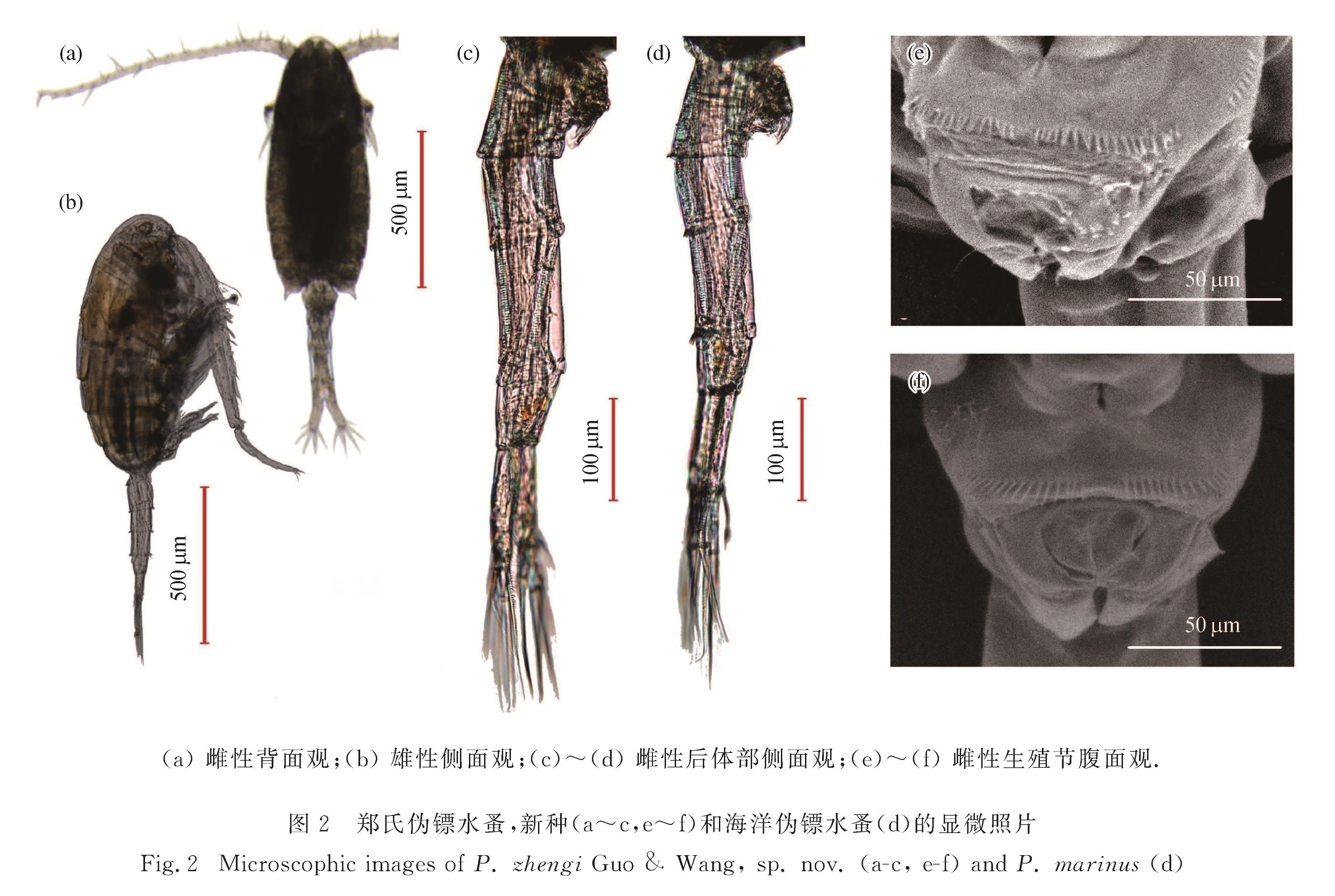
记述了厦门湾的伪镖水蚤科(Pseudodiaptomidae G O Sars, 1902)1个新种——郑氏伪镖水蚤(Pseudodiaptomus zhengi Guo & Wang, sp. nov.).形态上,新种雌性腹部各节光滑无小刺、生殖节的侧前端和生殖厣的前端各具1排长刺,与海洋伪镖水蚤(P. marinus Sato, 1913)存在明显的形态差异; 两者基于线粒体细胞色素氧化酶Ⅰ(mtCOⅠ)基因序列的种间遗传差异为22.4%~23.7%.模式标本(2016年4月至2017年7月采集于厦门湾)保存于厦门大学海洋科技博物馆.
A new species of Pseudodiaptomidae,Pseudodiaptomus zhengi Guo & Wang,sp.nov.,which was misidentified as P.marinus Sato,1913 by Zheng et al.(1965),is described.The differences between this new species and P.marinus are as follows: all female urosome segments without rows of coarse teeth on their distal margins,a row of spinules located on the anterolateral margin of female genital segment,and a row of spinules located in front of female genital operculum.The mitochondrial gene cytochrome oxidase subunit Ⅰ(mtCOⅠ)gene sequences support the conclusion based on morphological data,with distinct divergence values 22.4% to 23.7% between P.zhengi and P.marinus.All types of specimens,which were collected from Xiamen Bay during April 2016 to July 2017,are archived in the Museum of Marine Science and Technology,Xiamen University.