收稿日期:2018-08-12 录用日期:2018-10-24
< br/>基金项目:国家自然科学基金(U1805233,U1205123,31172438); 福建省自然科学基金(2018J01452)
< br/>通信作者:wshuang@jmu.edu.cn < br/>
< br/>通信作者:wshuang@jmu.edu.cn < br/>
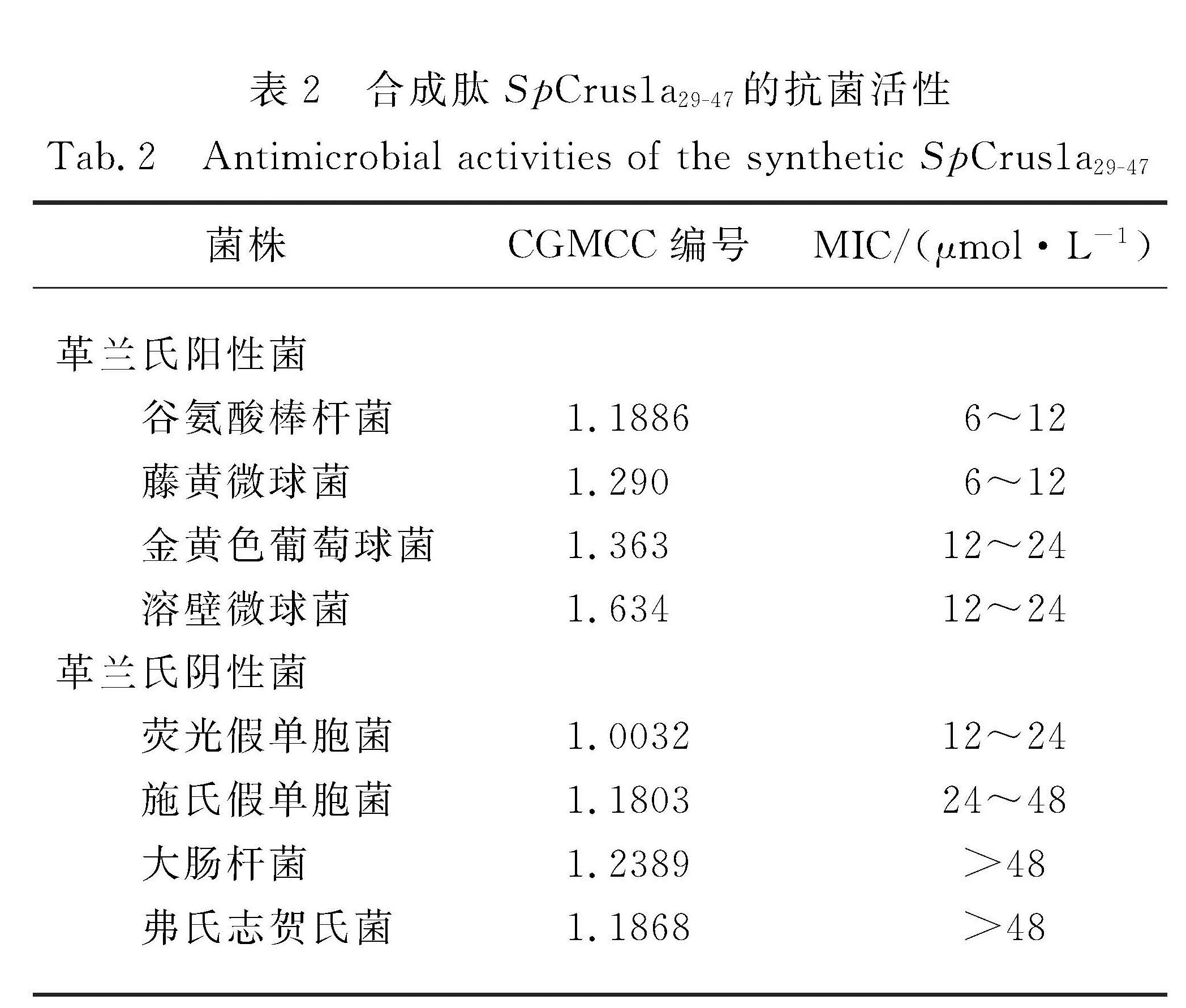
Crustins是一类较早发现并广泛分布在甲壳动物中且富含半胱氨酸的阳离子抗菌肽,能够参与抗菌免疫应答.从拟穴青蟹(Scylla paramamosain)中克隆获得一个Crustin基因新变体,命名为SpCrus1a,其cDNA序列全长744 bp,开放阅读框编码113个氨基酸,成熟肽分子质量10.03 ku,理论等电点8.30.表达分析结果发现其转录本主要存在于鳃、卵巢、上皮组织中,经脂多糖刺激后SpCrus1a表达会上调.体外合成SpCrus1a的第29~47位氨基酸(SpCrus1a29-47)进行抗菌活性实验,发现其对革兰氏阳性菌具有较强的抗菌活性,而对被测的革兰氏阴性菌抗菌活性较弱或无抗菌活性.浓度24 μmol/L的合成肽SpCrus1a29-47能够在5 min内杀死大多数的金黄色葡萄球菌(Staphylococcus aureus),在120 min内杀死全部的金黄色葡萄球菌.扫描电镜分析发现合成肽SpCrus1a29-47可造成金黄色葡萄球菌表面结构崎岖不平,高浓度SpCrus1a29-47会引起细菌大量死亡.上述结果表明SpCrus1a是抗菌肽Crustin的新变体.
Crustins are cationic cysteine-rich antimicrobial peptides discovered earlier and widely distributed across the crustaceans,and involved in the antimicrobial immune responses.In this study,a novel Crustin isoform was cloned from Scylla paramamosain,namely SpCrus1a.The full-length cDNA of SpCrus1a was 744 bp,with an open reading frame encoding a peptide of 113 amino acids.The predicted molecular mass of the mature protein was 10.03 ku with an estimated pI of 8.30.The transcriptional transcript of SpCrus1a was highly expressed in gills,ovary and carapace by RT-PCR analysis,and was up-regulated in gills after lipopolysaccharide challenge.The antimicrobial activity experiment was performed with the synthetic peptide from 29 to 47 amino acid residues of SpCrus1a(SpCrus1a29-47).It was found that the synthetic peptide showed more strong anti-bacterial activity against Gram-positive bacteria,but had no or little antimicrobial activity against selected Gram-negative bacteria.Most of Staphylococcus aureus were killed by the synthetic peptide SpCrus1a29-47(24 μmol/L)within 5 min and all were killed within 120 min.Observation by scanning electron microscopy revealed that the surface of S. aureus cells treated with the synthetic peptide SpCrus1a29-47appeared to be rough and distorted,and higher concentration of SpCrus1a29-47 could cause cell death.The above results demonstrate that SpCrus1a is a novel Crustin isoform.