收稿日期:2018-05-27 录用日期:2018-11-06
基金项目: 国家重点研发计划(2016YFA0601202); 国家自然科学基金(41776116)
通信作者:senjie.lin@xmu.edu.cn
基金项目: 国家重点研发计划(2016YFA0601202); 国家自然科学基金(41776116)
通信作者:senjie.lin@xmu.edu.cn
(State Key Laboratory of Marine Environmental Science,College of Ocean and Earth Sciences,Xiamen University,Xiamen 361102,China)
phytoplankton; South China Sea; community structure; environmental regulatory factor
DOI: 10.6043/j.issn.0438-0479.201805038
作为西太平洋最大的边缘海,南海位于热带-亚热带区域,其水文理化环境复杂,高度多样的浮游植物群落在丰度和生产力上均存在显著差异.迄今为止,人们对该海域浮游植物的时空分布及其影响的认识仍有限.通过收集历史资料,特别是近20年的研究成果,从浮游植物时空分布、环境调控因子以及受全球气候变化的影响三方面进行分析,总结南海浮游植物生态学研究的重要进展,进而针对研究现状的不完善之处探讨未来的研究方向,突出有待深入研究的领域,以促使南海浮游植物生态学的研究赶上世界同类海域的研究脚步.
Located in the tropical-subtropical region, the South China Sea is the largest marginal sea in the western Pasific Ocean. Due to the complexity of the hydrological,physical as well as chemical environment of the South China Sea ecosystem,significant variations exist in the abundance and productivity of the highly diverse phytoplankton community.So far,our understanding is still limited about the spatial and temporal distribution of phytoplankton and its regulating mechanism in this area.In this review,we gathered historical data,especially research results of the recent 20 years,summarizing and analyzing them in three perspectives,spatiotemporal distribution,environmental regulatory factor and the impact of global climate change.We emphasized the advances in the research methodology and important progresses in understanding phytoplankton ecology in the South China Sea.In addition,we proposed future research directions and highlighted areas where efforts should be invested to bring the South China Sea phytoplankton research up to speed with the equivalent ocean basins in the world.
南海又称为中国南海或南中国海,位于太平洋西北部,是由9个国家环绕的半封闭海域.南海北起北纬23°37',南至南纬3°10',东西跨越23°,总面积超过330万km2[1].广阔的南海海域蕴含着丰富的自然资源,浮游植物种类繁多,海洋鸟类大量繁衍,海洋鱼类分布广泛且极具经济价值,拥有大量的岛屿资源及丰富的海洋油气资源,具有“第二个波斯湾”之称[2].丰富的自然资源和特殊的地理环境使南海具有独特而复杂的海洋生态环境.
作为南海海洋生态系统中最为重要的初级生产者,浮游植物不仅是海水中溶解氧的主要供应者,同时也是海洋碳氮流通的主要贡献者,是鱼、虾、贝类等海洋经济动物的直接或间接食物来源[3].浮游植物对所在海域的生产能力起着重要作用,故有“海洋牧草”之称.据推算,海洋浮游植物通过光合作用产生有机碳的总量是高等植物的7倍左右,对CO2的吸收量则约为全球年产量的一半[4],因此南海浮游植物群落结构的稳定性与南海海洋生态系统的稳定性密切相关.浮游植物的组成结构和物种丰度变化直接影响海水水质[5]、生态系统物质流、能量流和生物资源的变动[6].因此只有充分了解南海浮游植物生态系统的历史、现状及未来的发展趋势,才能更好地利用南海资源,在人与自然和谐相处的同时实现对海洋资源的合理利用.本文聚焦近20年的研究动态,对南海浮游植物的分布特点、丰度变化及其影响因素等研究成果进行归纳总结,借此了解南海浮游植物生态现状及其在全球气候变化背景下的响应和反馈.
从20世纪80年代开始,国内海洋科研院所和高校等科研机构利用显微鉴定技术[7]、生物化学标记法[8]、流式细胞术[9]、系统发育法[10]等多种手段对南海浮游植物生态现状进行了多次有针对性的调查.通过总结近20年的南海浮游植物调查结果,发现南海浮游植物的物种组成和丰度分布具有明显的季节性和区域性[11].
1998年6月至1999年8月,国家海洋局第二研究所等单位对南海大尺度海域(5°~25° N,105°~120° E)进行了3次大规模航次调查,结果显示:南海海域表层的浮游植物共有5门54属119种,对比发现冬季的南海浮游植物细胞丰度(平均为8.460×103 L-1)明显高于夏季(平均为0.830×103 L-1),平均值达到夏季的10倍以上; 冬夏两季的浮游植物优势类群大致相同,均依次为硅藻、蓝藻和甲藻,其中硅藻在冬夏两季的细胞丰度占比分别达到95.8%和75.7%,且在冬季的绝对丰度和相对丰度均高于夏季,而蓝藻和甲藻在冬季的绝对丰度高于夏季,相对丰度低于夏季[12].中国科学院海洋研究所等单位于2004年冬夏两季利用Utermöhl方法对南海浮游植物群落结构进行了大规模调查研究[13],结果显示:冬季南海北部0~200 m海水中共发现浮游植物5门106属198种,细胞丰度为206.404×103 L-1,夏季共鉴定出浮游植物4门74属159种,平均细胞丰度为115.000×103 L-1; 对比发现南海北部海域冬季浮游植物的平均细胞丰度高于夏季,约为夏季的2倍,且2次调查期内南海浮游植物均以硅藻为主,其后依次为甲藻、金藻和蓝藻等.2009年至2010年的冬夏两季浮游植物群落结构调查结果[14]表明:夏季南海海域共鉴定出浮游植物4门72属150种,平均细胞丰度为26.49×103 L-1,冬季样品中共鉴定出浮游植物4门58属168种,平均细胞丰度为2.69×103 L-1; 与夏季相比,冬季浮游植物的平均细胞丰度偏低,但物种丰度却略高,浮游植物群落优势物种依次分属于硅藻门、甲藻门、金藻门和蓝藻门.Xiao等[11]利用13种色素标记南海浮游植物的9大类群(甲藻、硅藻、定鞭藻6型、定鞭藻8型、绿藻、隐藻、原绿球藻、聚球藻、青绿藻),经过12年的大规模调查总结了南海浮游植物色素的变动情况,结果显示:陆架区的浮游植物群落组成具有明显的季节性变化,具体表现为夏季浮游植物以硅藻为主,春季以原绿球藻和聚球藻为主,秋冬季以鞭毛藻和青绿藻为主; 反映浮游植物生物量的叶绿素a平均质量浓度也具有明显的季节性变化,由高到低依次为冬季(0.52±0.79)mg/m3>夏季(0.42±0.61)mg/m3>春季(0.25±0.32)mg/m3>秋季(0.22±0.31)mg/m3.综合1998年以来的多次冬夏两季调查资料(表1),发现南海浮游植物的物种组成和丰度分布具有明显的季节性,冬季的南海浮游植物细胞丰度明显高于夏季,春秋两季略低,冬季的物种丰度也较夏季略高,主要优势物种为硅藻,其后依次为甲藻、金藻和蓝藻等.个别航次调查结果差异较大,可能是由于调查区域及统计鉴定方法有所不同.
Ke等[18]的调查发现南海海域浮游植物的垂直分布基本趋势为细胞丰度随水深的增加而降低,在近岸及水深小于200 m的海域,总浮游植物及优势种硅藻的数量随水深的增加而减少,在水深10 m处浮游植物细胞丰度出现最大值,平均为43.38×103 L-1.而在水深大于200 m的深海区,真光层深度较大,表层浮游植物受强光的抑制以及营养盐的限制,细胞丰度在0~75 m随水深的增加而增加,大于75 m后则随水深的增加而逐渐减少,浮游植物细胞丰度最大值多出现在次表层(水深50~75 m)[3].通过分析南海浮游植物总叶绿素a的垂直分布,发现在垂直方向上的分布具有季节性,春季叶绿素a浓度最大值层在水深75 m左右,夏秋季的最大值层位于水深50 m处,冬季表层混合层的叶绿素a分布较为均匀,且随着混合层深度的增加而降低[11,19].
南海海域表层水体中浮游植物的分布受不同水文状况的影响和制约,表现出典型的区域分布特征,南海浮游植物数量或叶绿素等生物标志物总量的高值主要出现在沿海岸、河口外围、上升流区域[20],其他区域浮游植物细胞丰度的平面分布从沿岸向外海迅速减少.在富营养的沿海地区和大陆架上,聚球藻和微微型真核生物最为丰富,而在大陆坡和开阔的海洋中,原绿球藻最为丰富[21].针对南海北部海域浮游植物的平面分布情况,Ke等[18]的调查发现南海北部的广东沿海岸和东北部的台湾浅滩海域浮游植物细胞丰度明显高于平均水平.珠江淡水的输入为珠江口外围带来了丰富的营养盐,造成该海域表层海水具有低盐度、高营养盐的特性,使浮游植物生长不受营养盐限制,因此浮游植物细胞丰度较高; 而位于南海东北部的台湾浅滩海域由于受常年的上升流及地形影响,深层富营养盐不断上涌,有助于浮游植物的大量繁殖.这与Le等[3]在2004年的调查结果一致,李丽等[22]在2008年通过测定脂类等浮游植物生物标志物的表层分布情况也印证了该结果.位于南海西北部的西沙群岛海域浮游植物群落中种类最多的是硅藻,其次是甲藻和蓝藻,数量上以红海束毛藻(Trichodesmium erythraeum)为优势物种,大洋暖水高盐性的浮游植物类群所占比例相对较大,与沿海近岸浮游植物种类组成结构有一定区别[23]; 而位于西沙群岛以南的南海西部海域,浮游植物群落结构则以定鞭金藻、聚球藻和原绿球藻为优势类群[24].在南海南部南沙群岛海域的多次调查中,发现浮游植物细胞丰度的差异较大,但其分布情况的总体趋势相似,浮游植物细胞丰度高值区往往出现在南沙群岛的东部和南部,多次调查均显示该海域的浮游植物群落中甲藻种类数高于硅藻[25],暖水种和高盐种为南海南部海域的主要生态类型,细胞丰度达10%及以上的物种均为暖水种,且其中
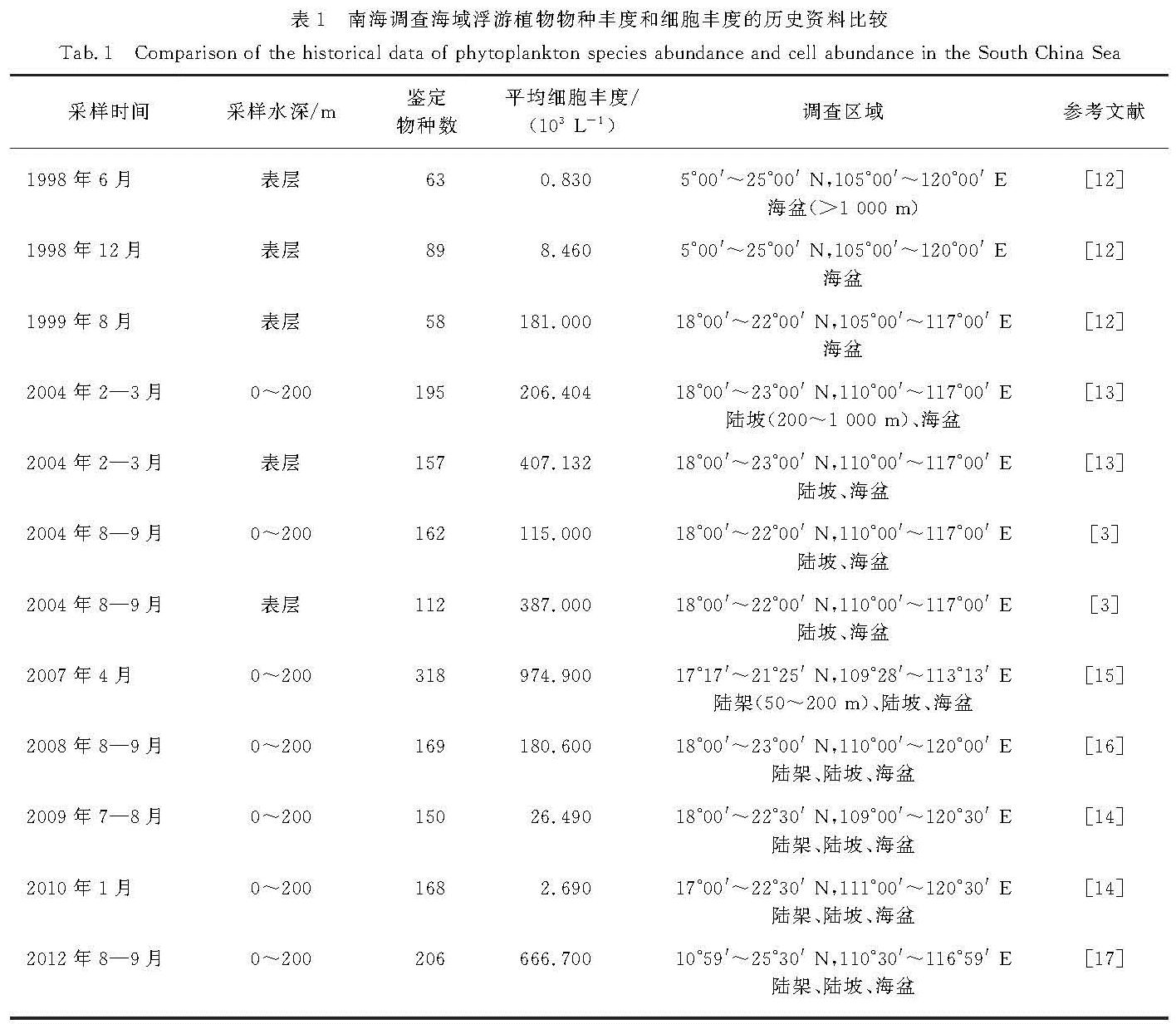
表1 南海调查海域浮游植物物种丰度和细胞丰度的历史资料比较
Tab.1 Comparison of the historical data of phytoplankton species abundance and cell abundance in the South China Sea
70%以上为高盐种[26].作为一个半封闭海,南海海域纬度跨度较大,局部理化特征复杂,浮游植物平面分布差异很大,因此需要更多的大规模调查才能更好地掌握其分布情况及规律.
南海广阔的面积和复杂的理化条件形成了其局部海域千差万别的水文环境,因而影响着局部海域的浮游植物群落结构乃至整个海洋生态系统的稳定性.影响南海海洋生态系统的因素主要有物理、化学、生物和地质4个方面,其中既包含营养盐分布等对海洋浮游植物影响较大的因素,又有海水pH值变动等影响微弱的因素.
温度是影响浮游植物光合作用、呼吸作用的重要因素,通常通过影响细胞代谢活动相关酶的效率而影响浮游植物的生物活动效率[27].绝大多数浮游植物的最适温度范围为20~30 ℃,一旦温度超过30 ℃,藻类各种生理活动均会受到一定程度的抑制,导致生长受到限制.南海海域表层水温受季节影响变化明显,受太阳直射点及太阳辐射影响,夏季南海整体海域表层水温约为28~29 ℃,温差在1 ℃以内波动,整体水温均一,对南海浮游植物分布的影响较弱[28]; 而冬季南海表层水温从北到南由22 ℃增加至28 ℃,温差较大,对浮游植物分布差异影响较大.冬季的温度更适宜南海藻类的生长,浮游植物生物量远高于夏季,温度可能是造成南海浮游植物群落结构及细胞丰度的季节性差异的主要原因.Tang等[29]利用卫星遥感技术对寡营养盐的吕宋海峡西南部海域的藻华现象进行了监测,发现该区域出现藻华现象时水温通常为一年中的最低值,Le等[3]在南海北部的研究同样发现了浮游植物种类丰度、生物量与水温之间的这种显著负相关关系.结合SeaWiFS卫星遥感对测量南海海域叶绿素a浓度及TRMM微波遥感对海表温度的分析,林丽茹等[30]发现普遍较高的叶绿素a浓度同较低的水温分布相对应,南海浮游植物生物量同水温存在较好的负相关关系.
盐度对浮游植物群落组成及细胞丰度都有一定的影响,作为优势类群的硅藻更适宜低盐度海水,而甲藻和蓝藻等对高盐度具有较高的耐受性; 南海海域盐度范围为24.19~34.59,由近岸到外海随着海水盐度受河流等淡水影响的减弱,盐度逐渐增加,浮游植物中硅藻的细胞丰度逐渐降低,甲藻和蓝藻等的细胞丰度升高,总体表现为当盐度大于30时,浮游植物生物量与盐度呈负相关[31].综上,南海浮游植物生物量与水温和盐度总体呈负相关,局部海域由于受上升流等其他因素影响,与水温的相关性较低.
营养盐是海洋浮游植物生长繁衍的基础物质,也是海洋初级生产力水平的主要调控因子,南海浮游植物的分布情况主要受营养盐影响,两者存在明显的线性相关性[31].作为典型的贫营养盐海域,南海表层营养盐的主要来源有陆源河流输入、深层海水上涌、随风尘输入和自身循环4种方式.受近岸人为因素、河流地理因素、季风变化因素等的影响,南海海水营养盐浓度总体呈现由近岸到外海逐渐降低的趋势[32],南北部浓度差异不大.
Wu等[33]通过总结近30年以来南海北部大亚湾沿海的浮游植物群落结构与营养盐调查数据,发现溶解无机氮的浓度显著增加,磷酸盐浓度则受人为影响而急剧下降,南海北部大亚湾海域浮游植物的限制因素已经从氮限制转变为磷酸盐限制; 尽管硅藻在浮游植物群落中仍然占主导地位,但占主导地位的物种却略有变化,同时由于营养结构的变化,网采浮游植物群落有逐渐微型化的趋势.对西沙群岛生态监控区的检测同样发现该海域氮磷元素的摩尔比大于16:1,磷是浮游植物营养盐限制的主要因子,其次为硅[23].魏玉秋等[34]对南海北部海域的调查发现硝酸盐、硅酸盐等具有明显的垂直层化现象,影响了浮游植物的垂直分布.彭欣等[31]通过对南海2个站位原始水样进行营养盐加富试验以研究营养盐对浮游植物的影响,发现当加富单一营养盐时浮游植物细胞丰度并无明显增加,同时加富氮、磷后浮游植物细胞丰度则达到培养前的5~20倍,而当同时加富氮、磷、硅后,原始水样中浮游植物细胞丰度总体显著增加,硅藻的细胞丰度大幅度增加且在浮游植物的总占比明显提高,说明该海域受多种营养盐限制,且营养盐加富不仅对浮游植物的增长有显著作用,同时也可以影响浮游植物的群落结构组成.硅元素是硅藻生长所必需的元素,其含量越高,硅藻总量和总占比越大,因此硅藻的分布趋势与海水中硅酸盐的分布趋势一致.有研究发现珠江口营养盐可被海流输送到盐度大于30的邻近外海,进而促进微微型浮游植物生长[35],表明河流携带的营养盐可能对邻近外海浮游植物产生影响.铁是束毛藻等固氮浮游植物固氮酶的必需元素,相对于其他浮游植物,固氮生物对铁的需求更大[36],因此铁元素也直接影响着束毛藻等在海洋中的分布.除了氮、磷、硅、铁等主要限制性营养盐外,最近有研究发现长期被认为是有毒金属的铝元素加富也会促进南海一些浮游植物种类的生长,其可能促进铁、磷的吸收并可结合超氧化物歧化酶,延缓生物物质降解[37].
光照是浮游植物进行光合作用的必需条件,直接影响着浮游植物的生物量.南海海域位于北半球低纬度区域,海水所受光辐射较强,平均光照时间较长.Cheah等[38]利用高光谱辐射计和快速重复频率荧光计对南海表面的光入射辐照度和叶绿素荧光参数进行了连续测量,发现南海浮游植物群落的生理条件主要受光照和营养盐浓度的影响.Sun等[39]研究发现部分近岸海区受河口等因素影响,虽然盐度低且营养盐丰富,但由于海水浑浊度高、透光性差,浮游植物细胞丰度不高,且近岸表层海水浮游植物细胞丰度高于次表层.陈兴群等[40]发现外海区域透光性好,表层海水的浮游植物受强光抑制,而次表层海水的光照强度更适宜浮游植物的光合作用,光照强度的差异造成外海区域次表层浮游植物细胞丰度高于表层,同时昼夜的光照变动影响了浮游植物的垂直运动.结合郝锵等[41]的调查数据和浮游植物细胞丰度的季节性变化,可知光照周期平均值与浮游植物细胞丰度呈负相关; 此外,通过对比南海北部冬夏两季初级生产力大小,发现冬季浮游植物光合作用速率高于夏季也与夏季光照强度过饱和从而造成光抑制有关.综上可见,光照对浮游植物的生物量及时空分布都有一定的影响.
季风对南海浮游植物生物量及分布的影响是通过海洋环流及上升流对海水中营养盐的影响实现的[42].大部分南海海域浮游植物生物量不高,且其季节性和空间分布主要受季风和海洋环境的影响[43],受海陆气压差或海气相互作用的触发可引起东亚季风在南海的爆发,包括盛行于夏季的西南季风和冬季的东北季风[44].在夏季,西南季风从5月中旬开始至9月结束,南海表层海水受风力搅动形成反气旋海流,在风力和海岸线的共同影响下,南海北部广东沿岸、海南岛东南部、越南东部和巽他陆架以北等沿海区域形成上升流,将深层营养盐带入表层与次表层海水,为浮游植物的生长提供大量的物质条件,进而提升了整个海域的生产力[43].对南海西南部的越南沿海地区的长期观测结果表明,在夏季西南季风的影响下,浮游植物特别是有害藻类大量繁殖[45-46],几乎每年都发生棕囊藻(Phaeocystis)的藻华现象,而当西南季风结束时,藻华也随之结束[47].在冬季,东北季风从11月开始至次年3月结束,其作用使南海表层海水与中层海水搅动形成混合层,同时在吕宋岛西北部和南部巽他陆架形成强烈的上升流,而中上层混合及上升流的形成均对海水表层的营养盐起到一定补充作用,进而促进浮游植物的生长[48].因此季风对南海的影响也体现在冬夏两季南海浮游植物生物量较高,并形成季节性波动.每年的3—6月往往是东北季风与西南季风的过渡时期,在此期间风向多变且模式混乱[49],不会在沿海或某一区域形成持续的上升流,同时由于没有西南季风的影响,南海反气旋的海洋环流在此时也是最弱的[50],因此季风过渡期的南海普遍表现出低营养盐水平.Voss等[51]发现,春季季风过渡期的浮游植物固氮率及其他水化学变量比夏季季风期低10倍.相对于冬夏两季的季风期,对于春季季风过渡期的调查研究相对较少,仍需更多的关注,从而更好地了解季风对浮游植物的影响.
南海沿岸上升流主要分布于南海东岸、广东东海岸、琼州海峡、雷州半岛东部、台湾海峡南岸及越南东南海岸[52],也有报道吕宋岛西北外海存在着季节性上升流[53].通常认为上升流是由表层辐散所致,夏季位于南海北部沿岸的上升流主要由西南季风驱动[54],而冬季位于吕宋岛西北则可能由附近的气旋环流汇合产生[55].不断上涌的上升流将低温而富营养盐的海水带到表层,提高了局部海域硝酸盐、亚硝酸盐等营养盐浓度[56],促进浮游植物的生长[57],形成一个与营养盐分布一致的浮游植物生物量和叶绿素a浓度高值区[56].与非上升流区域相比,上升流生态系统中经常可以观察到浮游植物的大量繁殖及藻华现象[58].南海沿岸上升流中浮游植物的主要优势种为硅藻,其次是甲藻和蓝藻.针对上升流中微微型浮游植物的研究发现沿海上升流水体中,真光层内的浮游植物群落以聚球藻为主,非上升流区真光层浮游生物群落则以原绿球藻为主[59].另有研究发现南海海域的上升流有时会发生断续[54],但其原因及对浮游植物的影响尚不明确,有待进一步研究.
在南海表层流速较大及海岸线或海底呈内凹形的海域常出现涡流,这些涡流分布广泛,北至湛江港外,南至南沙群岛东南,中尺度的涡流对生态系统的影响因其形成原因及冷暖类型等有所差异[60].这些涡流中浮游植物的群落结构[61-62]和生物量[63]等也与周围海水中的情况有所不同,暖涡和冷涡中的浮游植物主要种类为硅藻和甲藻[6],冷涡中的营养物质丰富,溶解氧含量低,叶绿素浓度及初级生产力都更高[6],而暖涡中的各种情况则刚好相反[64].Huang等[65]通过对南海的两个暖涡进行研究,发现暖涡会造成叶绿素浓度最大值层下移,暖涡中的浮游植物生物量较对照站位没有明显变化,但暖涡真光层中的浮游生物群落组成具有显著差异,因黑潮入侵形成的暖涡中优势种为原绿藻,而因地形原因形成的暖涡中优势种为定鞭藻.相对于冷涡而言,南海的暖涡情况更为复杂不定,因此对暖涡应予以更多的关注.
除营养盐、温度、光照等重要影响因素外,还有大气湿沉降、海底地形、黑潮水团等对浮游植物影响相对较小或仅对局部海域有明显影响的因素.Cui等[66]通过现场培养试验发现大气湿沉降可以提高海水中溶解无机氮与溶解无机磷的比值,雨水的输入不仅显著提高了叶绿素a浓度,还促使微微型浮游植物向微型浮游植物优势转移,从而改变了浮游植物群落结构.钱永甫等[67]通过分析南海模式海地地形发现其对海水质量输送和水温分布有重要影响,从而影响南海浮游植物的生物量及细胞丰度.Pan等[68]发现台风对南海浮游植物的生长及群落结构变化具有一定影响,超过一半的台风通过影响硝酸盐等营养盐的垂直向上运输而引起浮游植物的加速繁殖,进而提高海洋的初级生产力.黑潮在冬季入侵南海海域,与海水相互作用形成强烈的上升流,增强了营养盐的垂直向上运输,加快了浮游植物的生长繁殖,提高了初级生产力[69].由于黑潮水具有高温、高盐、低营养盐的特点,其入侵造成吕宋海峡附近海域近海性的硅藻细胞丰度减少,而远海性、暖水性硅藻和甲藻的细胞丰度则显著增加,影响了局部海域浮游植物的群落结构[70].浮游植物大多数体积微小,运动能力不强,无法自主地大范围移动,通常随海水流动而移动,因此水团对浮游植物的分布有直接影响.水团的锋面起到混合营养盐、富集浮游植物的作用,往往是生物活动较多的区域[70].不同水团的交融使水体强烈混合,直接影响上层海水中悬浮体物质组成和分布[71],进而影响局部的浮游植物生物量、群落结构和分布[42].综上可见,诸多因素造成了南海复杂的生态环境,也共同影响着南海浮游植物的发展变化.
全球气候变化对南海的影响主要体现在海水升温、海洋酸化、海平面升高、紫外线辐射加剧等方面.自1977年以来,近海岸浮游植物的生物量和群落结构在全球气候变化和人类活动的影响下已经发生了显著变化[72-73].全球气候变化是影响浮游植物群落结构变化的主要因素[72],Zhang等[74]通过分析近30年来南海表层温度变化趋势,发现南海表层温度几乎每10年增加0.194 ℃,浮游植物分布范围北扩[75].唐森铭等[76]通过总结遥感数据与资料分析了南海表层温度变化情况及对浮游植物的影响,发现南海浮游植物群落结构发生了显著变化,尤其是微微型浮游植物和甲藻,而暖水性浮游植物向北迁移,同时叶绿素a浓度减小,初级生产力降低,与水温呈负相关.唐启升等[77]研究发现酸化的海水中,颗石藻等海洋钙化生物的钙化速率降低,较大粒径的浮游植物比微微型浮游植物适应性更强,海水pH下降时浮游植物种类数出现明显下降,浮游植物群落结构变得较为单一.受气候变化的影响,从1970年开始,南海海域的海平面以平均每年(3.14±0.40)mm的速度上升,使南海由沿岸向陆地延伸扩张,带来沿岸海水环境的变动,造成沿岸生态系统的不稳定性[78].由全球气候变化引起的臭氧层受损及局部空洞使海洋表层受紫外线辐射加剧,造成部分浮游植物的垂直迁移[79]、核酸受损等现象,直接影响海洋上层浮游植物的群落结构与细胞丰度,对紫外辐射较为敏感的硅藻竞争力也相对下降,进而对局部生态系统造成影响[80].近50年来南海季风呈现出逐渐减弱的趋势,尤其是秋冬季的风速减弱明显,直接导致沿岸上升流减弱,影响营养盐的输送,降低了局部海域初级生产力[81].
随着对南海海洋科学研究的大力投入,近年来围绕南海各方面的研究都取得了很大程度的进展,特别是对海洋浮游植物的研究.由于各种科考航次的增加及研究设备和方法的不断改进,已经获得了一定的基础数据及资料积累,但是目前海洋浮游植物的研究范围及手段仍然受到很多限制,某些方面的研究仍然很欠缺,主要表现在以下3个方面:
1)调查方法传统,仍需改进完善.对南海浮游植物的研究已有数十年,但是应用于南海浮游植物调查的方法仍然以传统的显微镜鉴定为主,耗时费力,经验依赖性较强且人为误差较大,同时面临着人才匮乏的现象.尽管现在逐步采用了一些更简便快捷的方法,但仍存在不足之处,如利用色素等生物标志物鉴定难以精确到物种[11],DNA条形码鉴定数据库不全面而不能准确定量[82]等,各种方法需要不断改进、相互验证并加以完善; 遥感技术在分析南海叶绿素a浓度和初级生产力方面应用广泛,但目前的研究或为航次调查或为遥感调查,将航次调查与遥感调查同步结合互相参照的研究较少,使得很多研究不能深入开展,很多结论无法实际验证,因此需要克服局限性进行多学科综合性的调查.
2)研究目标单一,需要加强结合.目前对南海浮游植物的研究多为某特定条件下浮游植物群落组成及丰度分布的调查,在将海洋浮游植物与细菌、海洋浮游动物等其他生物结合的研究方面仍有很大空间.研究方法上,随着各种宏组学方法在环境生物研究方面的广泛应用,南海浮游植物调查结合宏转录组、蛋白组等组学分析的研究也有待进一步加强,从而在分子水平上探究机制、总结规律,这也是海洋生态研究的发展趋势.
3)缺乏长期监测,存在研究空白.关于全球气候变化对浮游植物的影响已有很多研究,但对南海海域浮游植物应对全球气候变化的研究并不多,且缺乏长期监测.浮游植物对气候变化的响应及其机制往往需要十年甚至数十年的观测才能总结出可靠的结论,而这方面的工作还处于起步阶段,急需建立类似于加勒比海的百慕大大西洋时间序列研究(BATS)和夏威夷海洋学时间序列(HOT)的长期研究站位.已有研究主要集中在东南亚时间序列观测(SEATS)站开展[83],但尚待完善系统的长期研究与监测平台.影响浮游植物的环境因素仍有很多研究空白之处,如:目前已经初步了解冬夏两季季风对南海浮游植物的影响,但季风过渡期的情况却并不清楚; 目前已经对南海海域的水团有了各种认识和划分,却并不清楚每个水团中浮游植物群落结构的异同.
随着研究方法的不断改进,南海浮游植物生态也将得到更全面深入的研究.首先,高通量测序因其高准确性、高灵敏度、高通量等优势,在南海浮游植物多样性的研究中应用广泛,测序技术的创新使得成本降低的同时通量及读长都不断提高,在目前基于18S和转录间隔区(ITS)测序[25]的基础上,逐步扩展到宏基因组测序,使得南海浮游植物多样性研究从物种(遗传)多样性迈向功能多样性.其次,DNA测序与RNA测序的结合促进了对浮游植物代谢活性的研究[84],浮游植物网络分析不断发展,促使针对南海浮游植物的研究由目前的多样性研究深入到物种间生态关系的剖析.再者,结合航次调研,遥感监测从总叶绿素水平向浮游植物类群及种类水平逐渐改进,有助于实现对南海初级生产力及浮游植物群落组成等的连续监测.此外,传感器与分子探针结合的改进将在很大程度上促进南海浮游植物多维时空实时监测的发展,浮游植物群落及种群动态计算机模型也将不断完善,对于了解南海水质、初级生产力、群落演替规律及海洋碳循环研究等都有重要意义.
目前我国的研究海域多数集中在南海北部(14° N以北),其他区域的各种基础数据及观测记录还比较缺乏.因此,在对南海调查研究中需要加强与周边国家的合作,以对南海区域开展更全面深入的研究,同时需要加强古今结合及不同研究方向的结合,充分了解在全球气候变化背景下浮游植物群落结构的演替机制,掌握南海浮游植物的演化趋势及物质和能量在食物链中的流动和传递规律,合理利用南海海洋资源,开发其巨大价值,实现人与自然的和谐相处.
