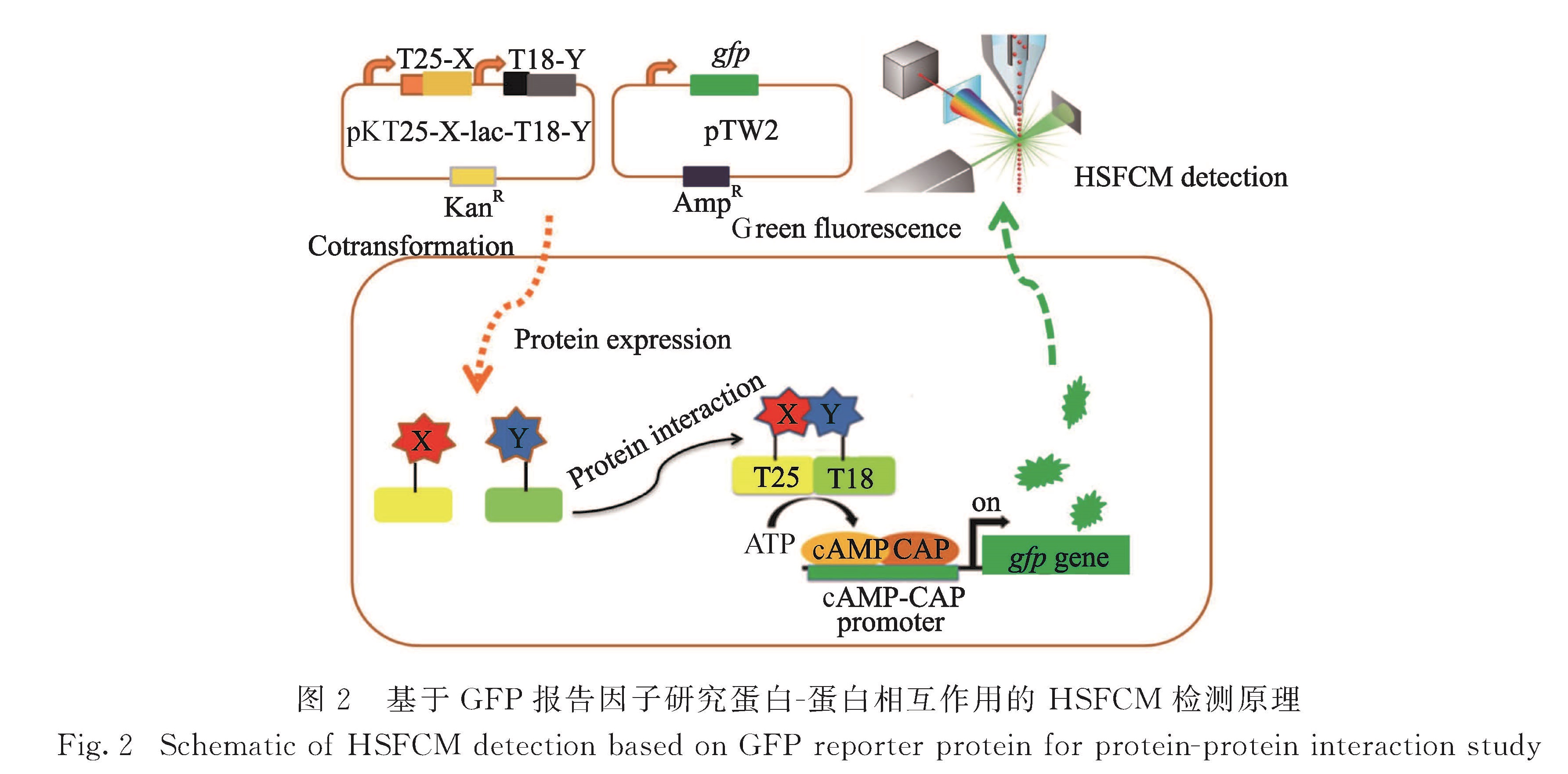
蛋白-蛋白相互作用是细胞生化反应网络的重要组成部分,参与了各种代谢途径和生理过程[1-2].对蛋白-蛋白相互作用进行分析有助于解析疾病的发生机制,对药物的设计筛选具有重要的指导意义[3].基于腺苷酸环化酶(adenylate cyclase,AC)功能重构的细菌双杂交(bacterial adenylate cyclase two-hybrid,BACTH)系统是蛋白-蛋白相互作用经典技术酵母双杂交系统的衍生,具有实验周期短、转化率高、蛋白无需核定位等优点[4].BACTH系统利用了百日咳博代杆菌(Bordetella pertussis)中的AC催化域是由两个互补片段T25和T18组成的特点,当T25和T18分别与可发生相互作用的多肽或者蛋白质融合表达时,嵌合蛋白的二聚化可以导致AC片段功能性的互补以及环腺苷酸(cyclic adenosine monophosphate,cAMP)的生成,cAMP与分解代谢激活蛋白(catabolite activator protein,CAP)结合,从而调控报告基因的表达[5].目前,BACTH系统已被广泛应用于特定蛋白对的相互作用鉴定以及蛋白质相互作用文库的筛选[6-7].在生命医学研究方面,BACTH系统也发挥了重要的作用.Zoued等[8]通过对细菌Ⅵ型分泌系统的收缩鞘中各组件的相互作用分析,揭示了其聚合的操纵机制以及内管装配的协调机制.Tao等[9]利用BACTH系统对肉毒毒素B进行饱和诱变筛选,增强了工程型肉毒毒素B特异性地靶向人类突触结合蛋白Ⅱ,提高了肉毒毒素B对神经性疾病的治疗效果.
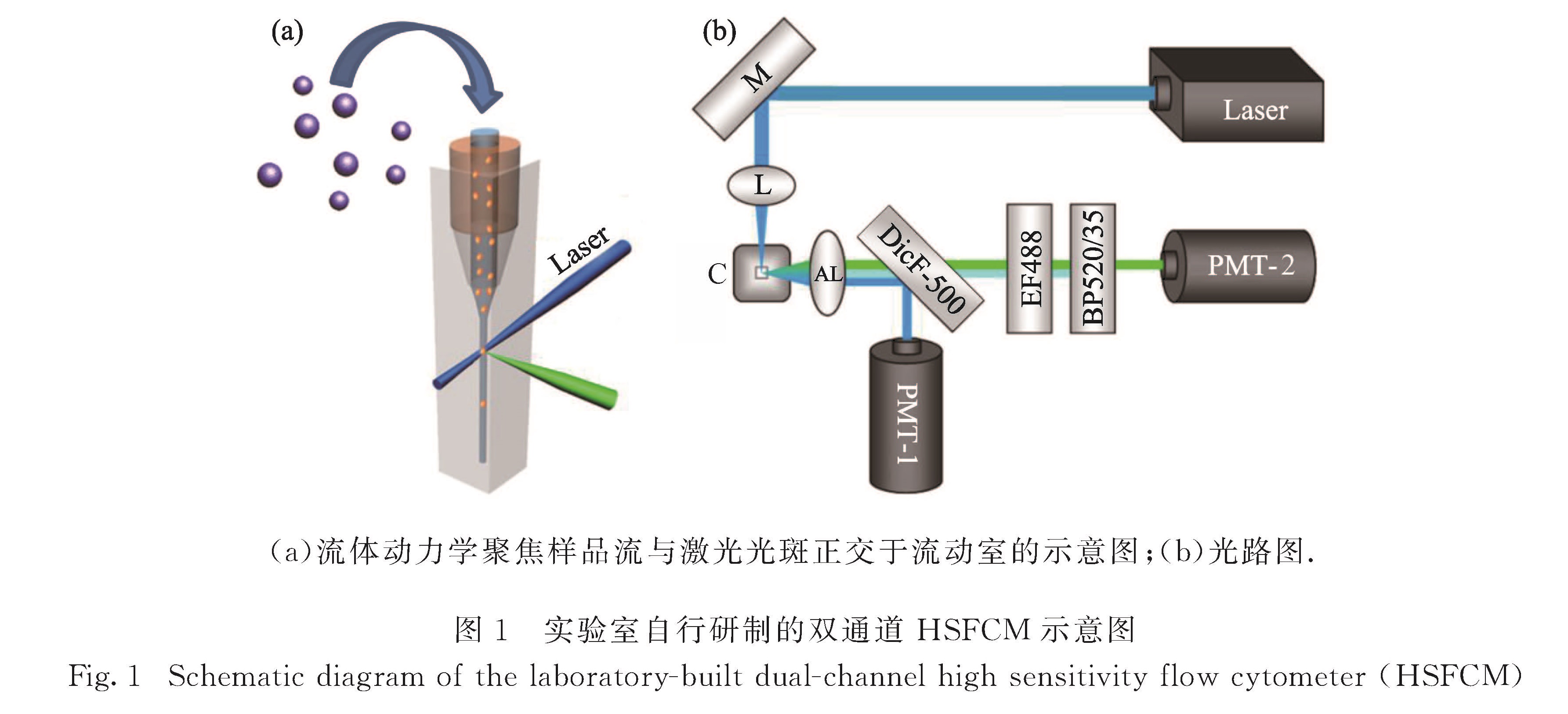
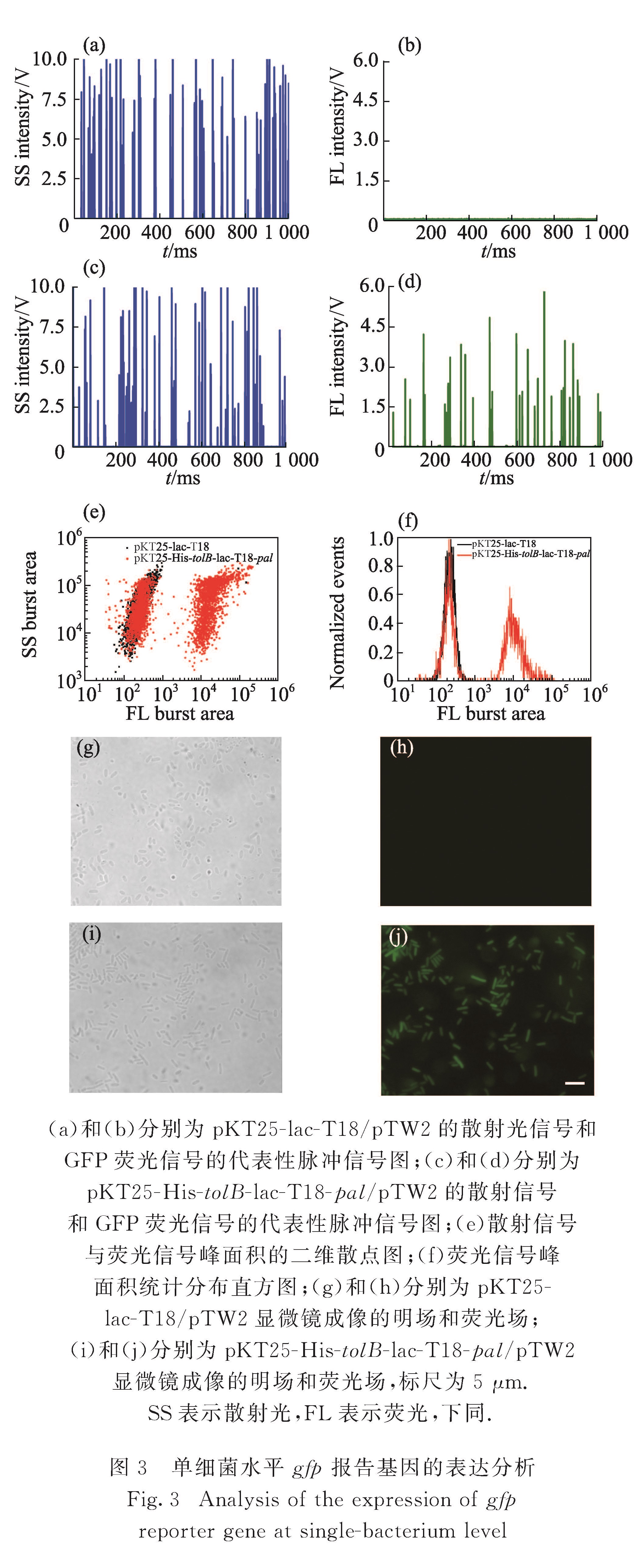
BACTH系统主要以lacZ为报告基因,对于报告基因所表达的β-半乳糖苷酶,传统的检测方法主要是通过测定邻-硝基酚-β-D-半乳糖苷或4-甲基伞形酮酰-β-D-吡喃半乳糖苷这类特异性底物水解产生的分子吸光度或荧光的变化对酶活性进行检测[4,10].然而,这类方法得到的是大量细菌中β-半乳糖苷酶表达量及活性的平均结果,掩盖了细菌个体间β-半乳糖苷酶表达量的差异.在单细菌水平考察β-半乳糖苷酶的表达有利于更好地解析基因表达的随机性、基因调控以及微生物的生理过程,对深入理解BACTH系统的工作机理从而针对性地改进实验方法也有极大帮助.此外,单细菌水平的蛋白-蛋白相互作用研究有助于更准确地考察蛋白在细菌介导的活细胞中相互作用的状况,对于揭示蛋白-蛋白相互作用相关信号转导的分子机制具有明显优势[11].流式细胞术是一种功能强大的单细胞分析手段,具有检测速度快、精度高和多参数分析等特点,已在蛋白质组学和系统生物学中发挥重要的作用[12].然而,由于细菌个体微小,传统的流式细胞仪难以在单细菌水平对其内部的蛋白质进行定量分析并考察蛋白-蛋白相互作用.结合瑞利散射和鞘流单分子荧光检测技术,本课题组自行研制的超高灵敏流式检测装置(HSFCM)散射光通道可实现粒径为24 nm的二氧化硅纳米颗粒和粒径为7 nm的金纳米颗粒的检测,荧光通道的检测灵敏度为3个Alexa Fluor 532分子[13],较传统流式细胞仪的散射和荧光检测灵敏度分别提升4~5个数量级和1~2个数量级,已被开发应用于细菌生化体系的分析[14-18].
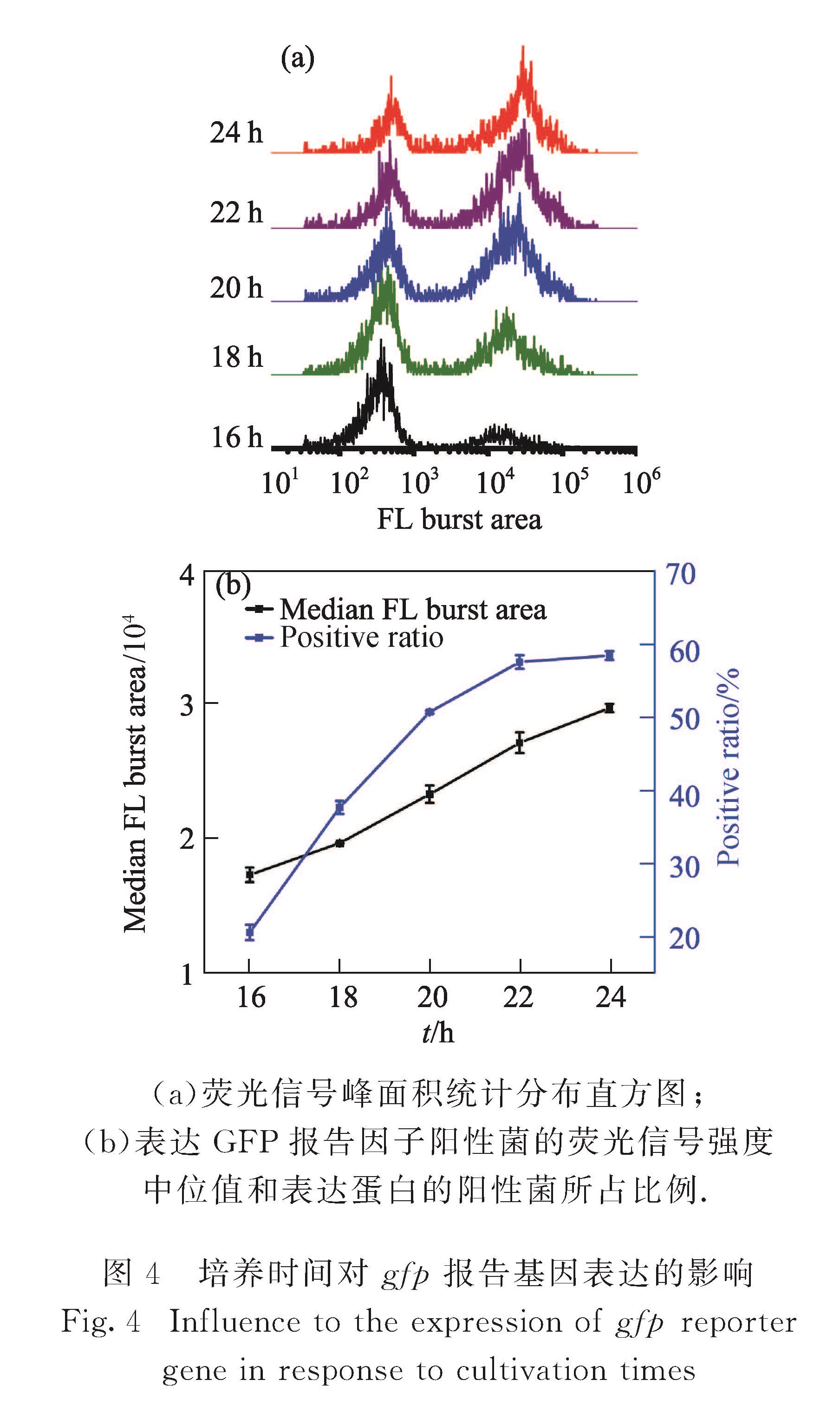
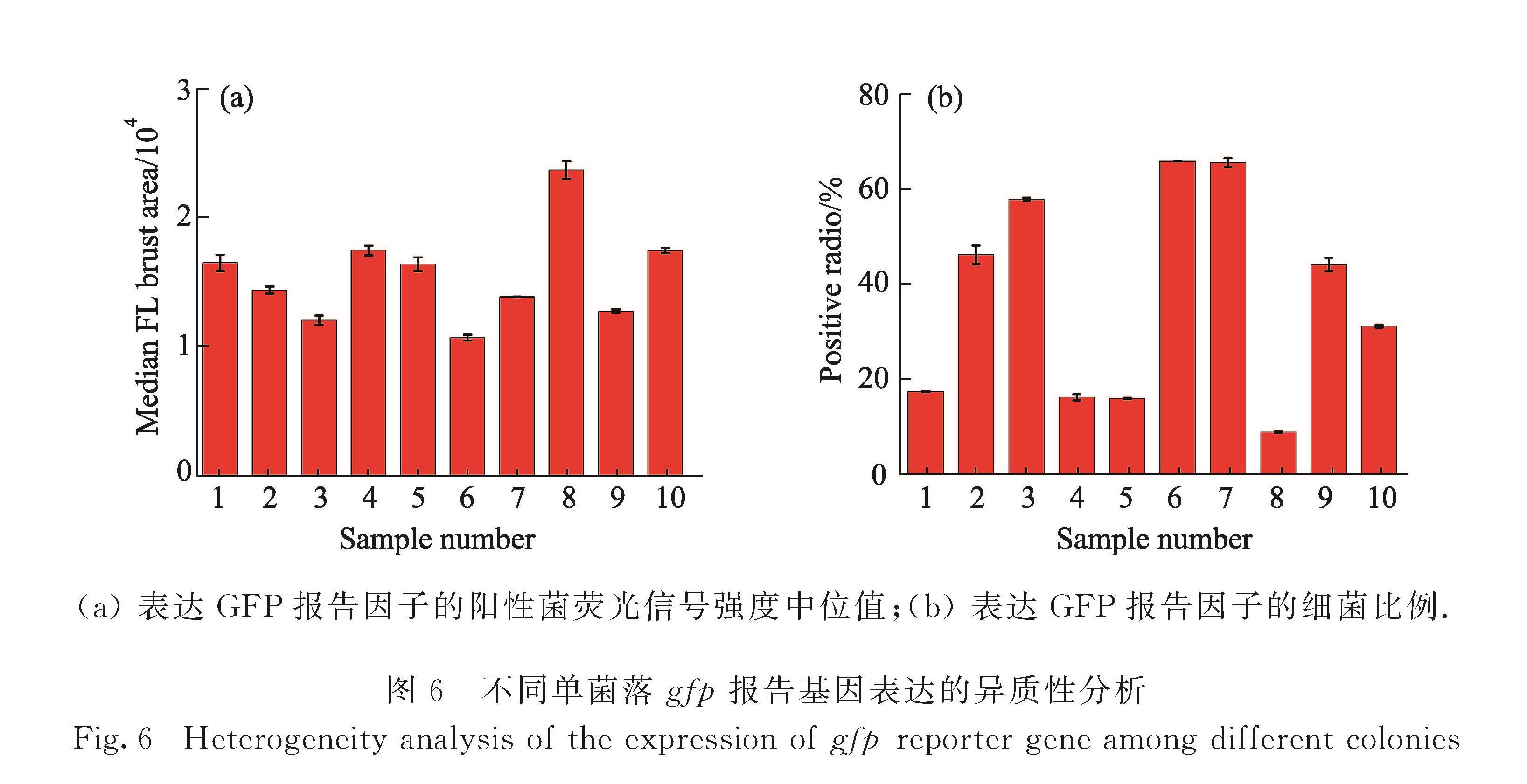
在本课题组的前期研究中,采用双色免疫荧光染色法对相互作用蛋白对和β-半乳糖苷酶同时进行荧光标记,并通过相对报告因子表达量实现了单细菌水平蛋白-蛋白相互作用的定量分析[19].然而免疫荧光标记需要对细菌进行固定、破膜和染色处理,操作相对复杂.发展一种简单、快速的蛋白-蛋白相互作用检测方法,将有效推进蛋白质相互作用研究.基于荧光蛋白的分子探针和标记方法已成为活细胞内研究生物大分子或细胞功能的重要手段.荧光蛋白能形成内部发色团,无需添加任何辅助因子,不需要对细胞进行处理,可直接对其荧光表征,实现天然状态下的细胞生化分析[20].本研究中引入lac启动子控制的gfp基因作为蛋白质相互作用的报告基因,采用实验室自行研制的HSFCM建立了一种单细菌水平的、简单、快速、高通量的蛋白-蛋白相互作用分析方法,在单细菌水平对GFP荧光信号强度和表达GFP报告因子的细菌比例分析,分别考察了培养时间和诱导剂浓度对gfp报告基因表达的影响,并对不同单菌落gfp报告基因表达异质性进行分析,以期为相互作用蛋白对的检测及筛选提供有效的分析手段.