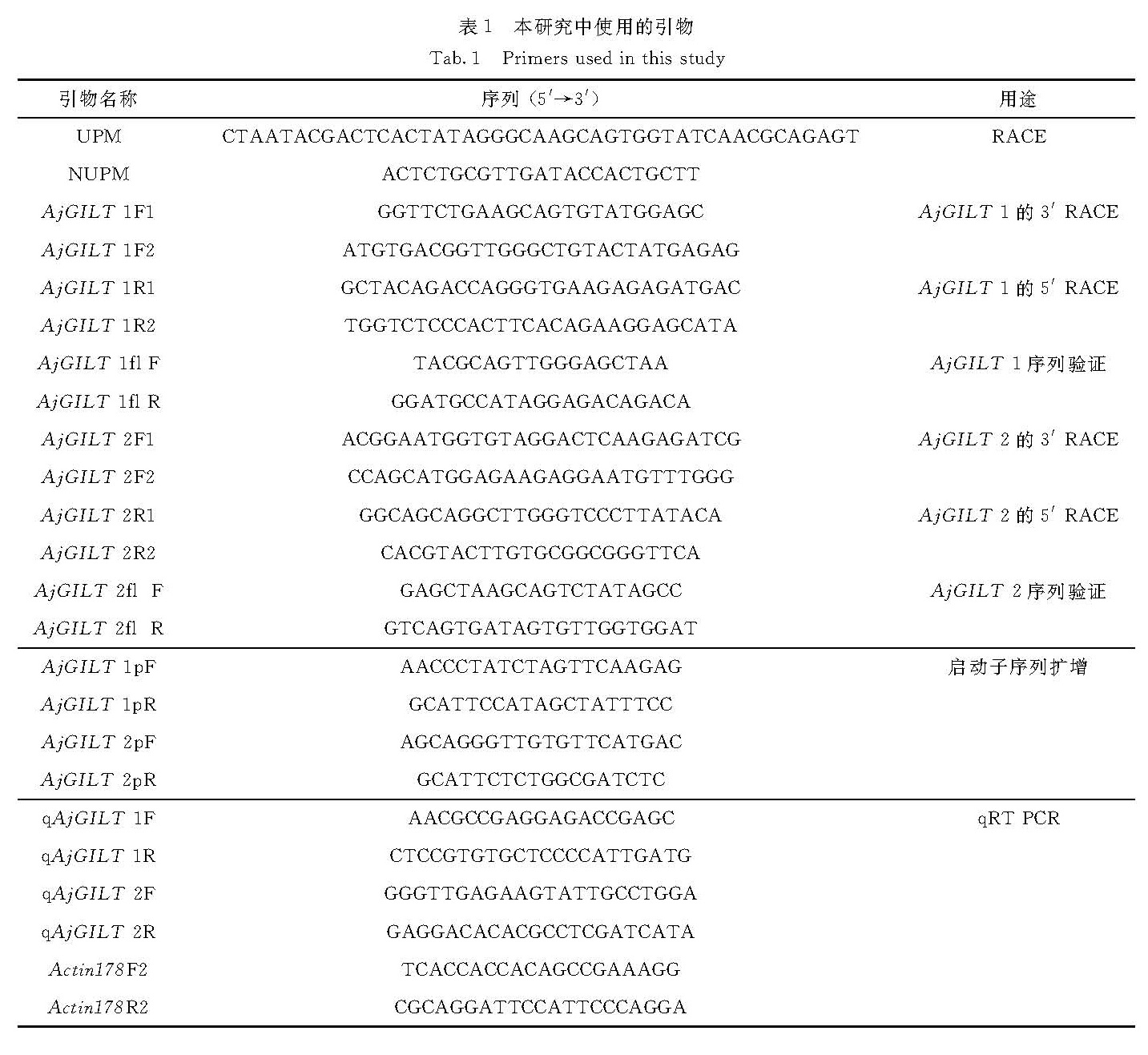
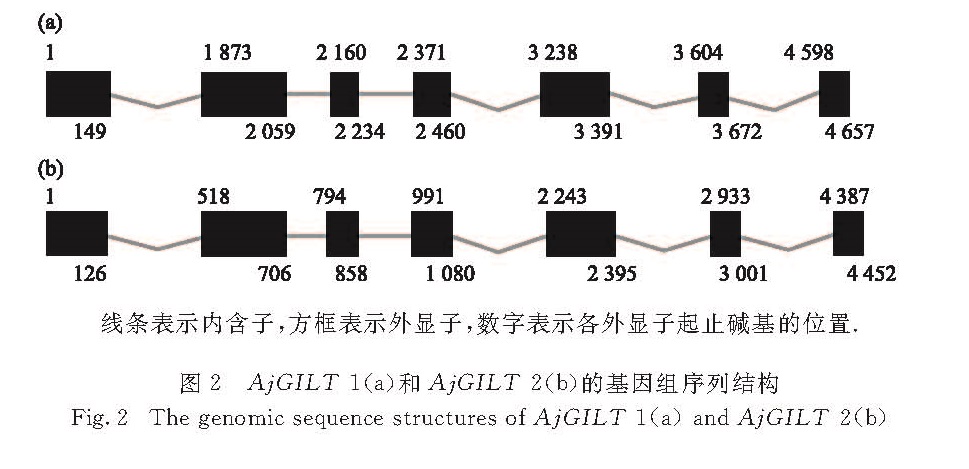
在日本鳗鲡(Anguilla japonica)中克隆获得2个γ-干扰素诱导的溶酶体巯基还原酶(GILT)基因AjGILT-1和AjGILT-2,并对其序列进行生物信息学分析,进而利用实时荧光定量PCR分别检测2个基因在不同组织器官中的表达水平,及其在脂多糖(LPS)、聚肌胞苷酸(PolyI:C)刺激和迟缓爱德华氏菌(Edwardsiella tarda)感染后的表达变化.结果显示:2个基因编码的蛋白序列中均含有GILT家族的特征序列CQHGX2ECX2NX4C、活性位点CXXC、2个糖基化位点和11个保守的半胱氨酸残基,基因组序列均含7个外显子和6个内含子.2个基因启动子中都存在多个转录因子结合位点,如γ-干扰素活化位点以及特化蛋白1结合位点; 不同的是AjGILT-1启动子区存在多个核因子-1、激活蛋白-1结合位点及干扰素刺激反应元件,而在AjGILT-2启动子区未发现.AjGILT-1在鳃和肠中表达量最高,而AjGILT-2在性腺、头肾和脾脏中表达量最高; 除肠道外,同一组织中AjGILT-2的表达量均高于AjGILT-1.LPS、PolyI:C刺激和E. tarda感染后,AjGILT-1与AjGILT-2的表达在鳃中仅有少数变化,而在头肾和中肾中则变化较明显.综上所述,AjGILT-1和AjGILT-2基因可能参与了日本鳗鲡受病毒和细菌感染后的免疫应答.
Two interferon-γ-inducible lysosomal thiol reductase(GILT)genes named AjGILT-1 and AjGILT-2 were cloned and identified from Anguilla japonica.The deduced AjGILT-1 and AjGILT-2 coding sequences contained the major characteristic features of the GILT protein family:the GILT signature sequence CQHGX2ECX2NX4C,the active site CXXC,two potential N-linked glycosylation sites and eleven conserved cysteines.The AjGILT-1 and AjGILT-2 contained seven exons separated by six introns.Moreover,AjGILT-1 and AjGILT-2 shared many transcription factor binding sites in the promoter region,such as interferon-γ activation site and serum response factor binding sites.The difference between them is that AjGILT-1 has nuclear factor-1,activator protein-1 binding sites and interferon stimulation reaction element,while AjGILT-2 does not.Quantitative real-time PCR analysis revealed that the expression level of AjGILT-1 was highest in gill and intestine,while the expression level of AjGILT-2 was highest in sex gonad,head kidney and spleen.The expression level of AjGILT-2 was higher than that of AjGILT-1 in most tissues,except for the intestine.After the stimulation with lipopolysaccharide(LPS),polyinosin-polycytidylic acid(PolyI:C)or infection with Edwardsiella tarda,there were only a few expression changes of AjGILT-1 and AjGILT-2 in the gills,while the expression changes in the head and middle kidneys were more obvious.In summary,AjGILT-1 and AjGILT-2 may be involved in immune responses to infection of viral and bacterial pathogens.