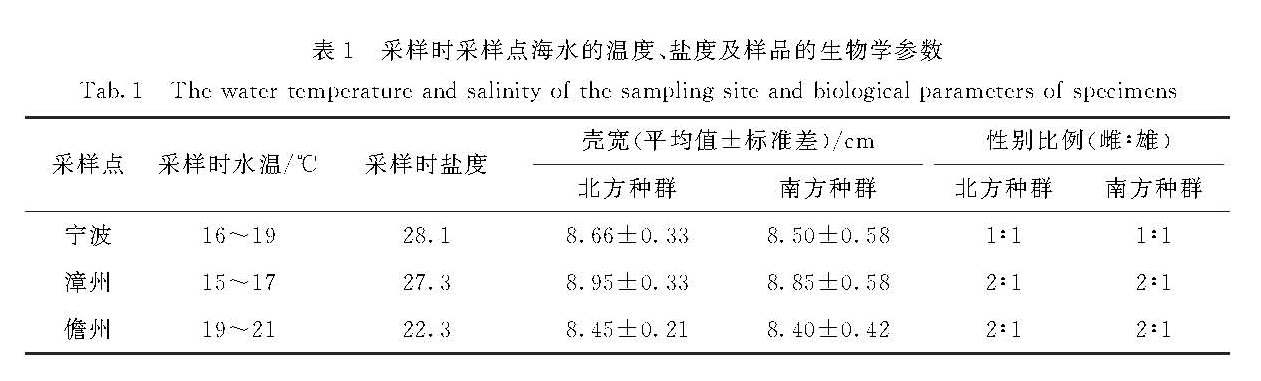
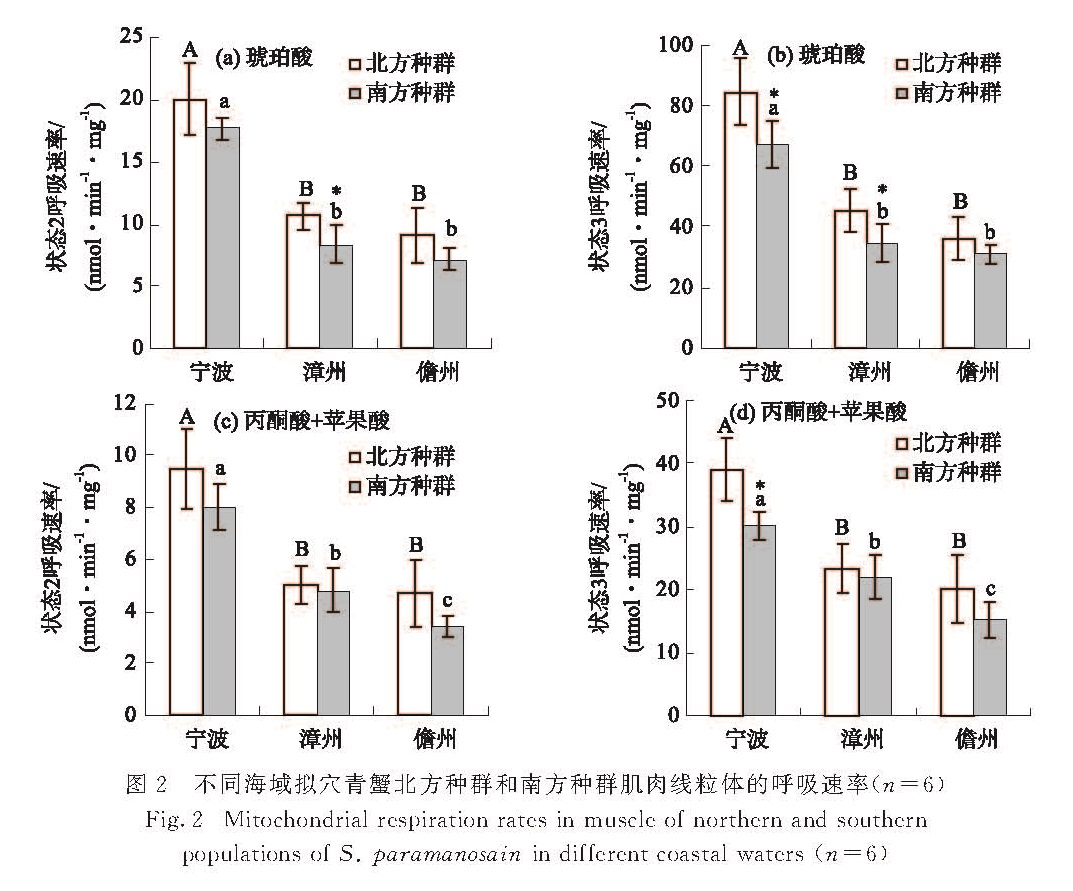
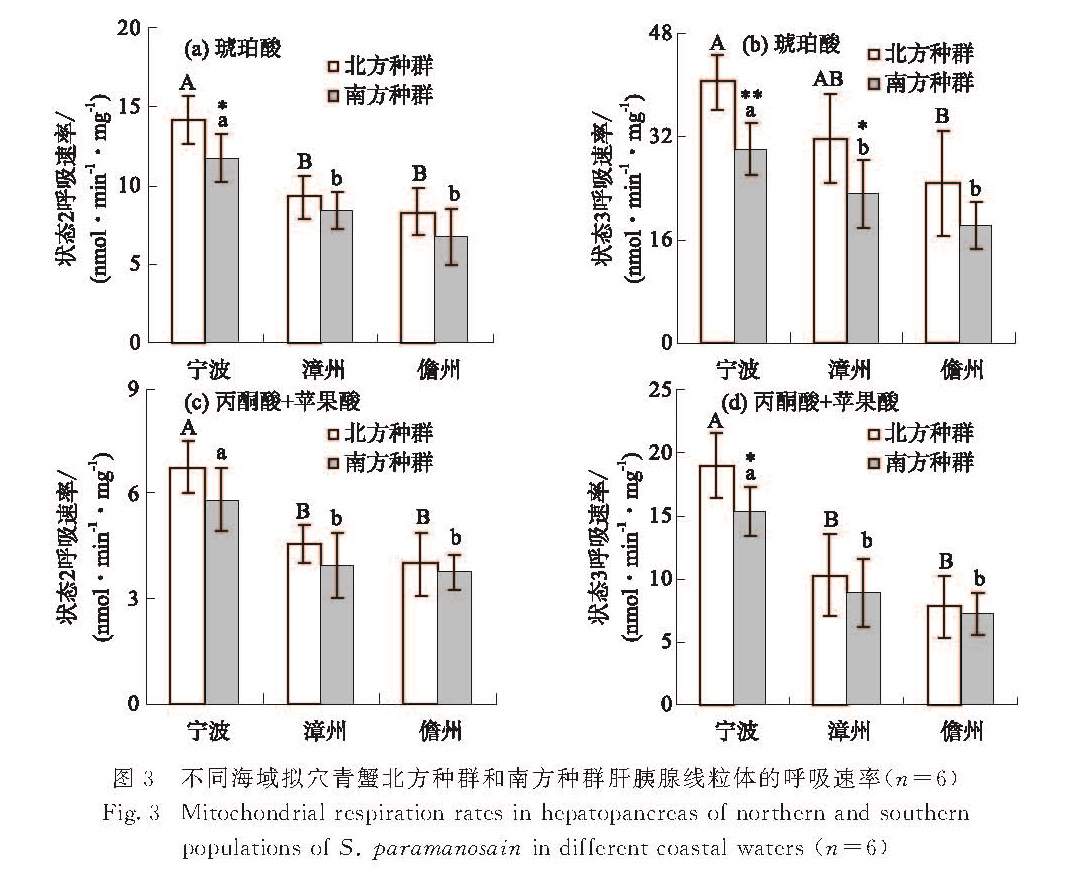
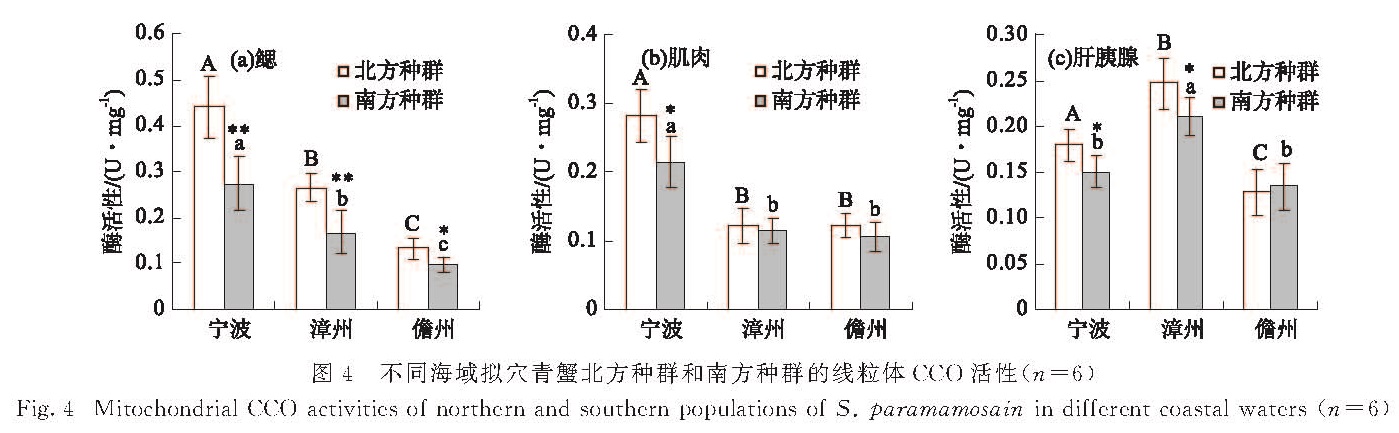
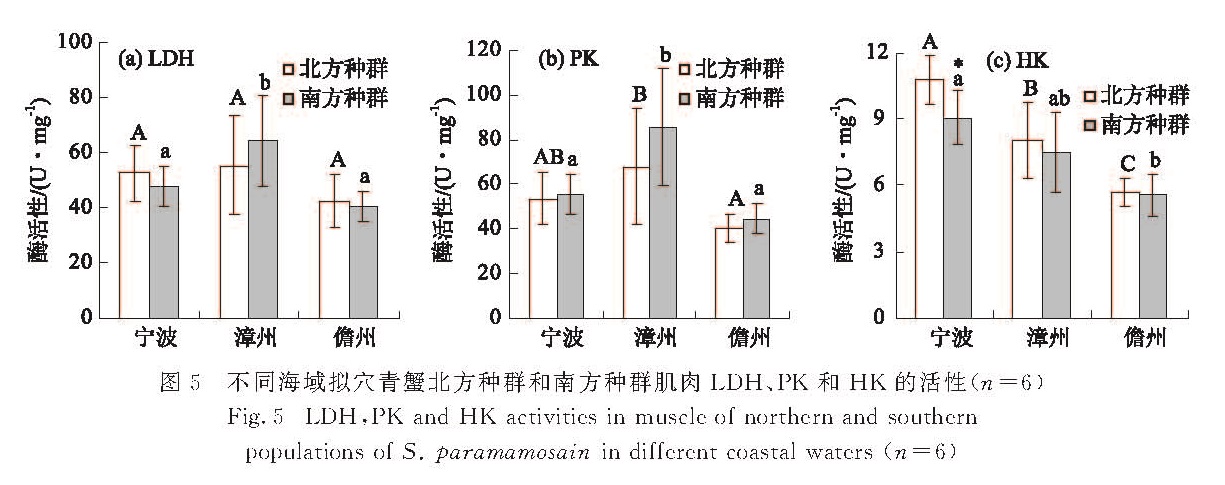
已有研究显示分布于我国东南沿海的拟穴青蟹(Scylla paramamosain)可能已分化为南、北2个种群.在低温季节从不同纬度的3个海域采集拟穴青蟹的野生个体,比较其南、北种群线粒体呼吸速率和酶活性的差异.结果如下:除了乳酸脱氢酶、丙酮酸激酶以及肝胰腺细胞色素C氧化酶外,无论是北方种群还是南方种群,生活在纬度较高的宁波海域的拟穴青蟹的线粒体呼吸速率和酶活性都显著高于生活在纬度较低的儋州海域的个体; 同一海域的南、北种群间,鳃、肌肉和肝胰腺线粒体呼吸速率和细胞色素C氧化酶活性总体上都是北方种群高于南方种群,纬度越高的海域的南、北种群间差异越显著; 南、北种群肌肉的乳酸脱氢酶、丙酮酸激酶和己糖激酶活性相比,宁波海域北方种群的己糖激酶活性显著高于该海域的南方种群.上述结果表明,分布于我国东南沿海的拟穴青蟹的北方种群比南方种群对温度有更有效的代谢补偿能力,且纬度越高,二者的差异越显著,由此可见分布于我国东南沿海的拟穴青蟹的北方种群比南方种群更能适应低温环境.
Recent research has identified that the mud crab,Scylla paramamosain,in the southeast coast of China can be sub-divi-ded into northern and southern populations.The present study measured and compared the mitochondrial respiration rates and enzyme activities of northern and southern populations of the mud crab,S. paramamosain,sampled during low temperature seasons and from three locations at different latitudes.It was shown that for both northern and southern populations,with the exception of lactate dehydrogenase,pyruvate kinase and cytochrome C oxidase in hepatopancreas,the mitochondrial respiration rates and enzyme activities of crabs collected from higher latitude Ningbo coastal waters were significantly higher than those from Danzhou coastal waters at lower latitude.As for northern and southern populations collected at the same location,the mitochondrial respiration rates and cytochrome C oxidase activities in the gill,muscle and hepatopancreas of crabs of the northern population were generally higher than those of the southern population from all three locations with the differences increasing with latitude.For lactate dehydrogenase,pyruvate kinase and hexokinase activities in muscle,hexokinase activities of crabs of the northern population were also significantly higher than those of the southern population collected from Ningbo coastal waters.The results of the present study indicated that the northern population of S. paramamosain had strong metabolic compensation capacity than the southern population under cold environments,especially at higher latitude,confirming the hypothesis that the northern population of S. paramamosain adapted better to low temperature than the southern population.