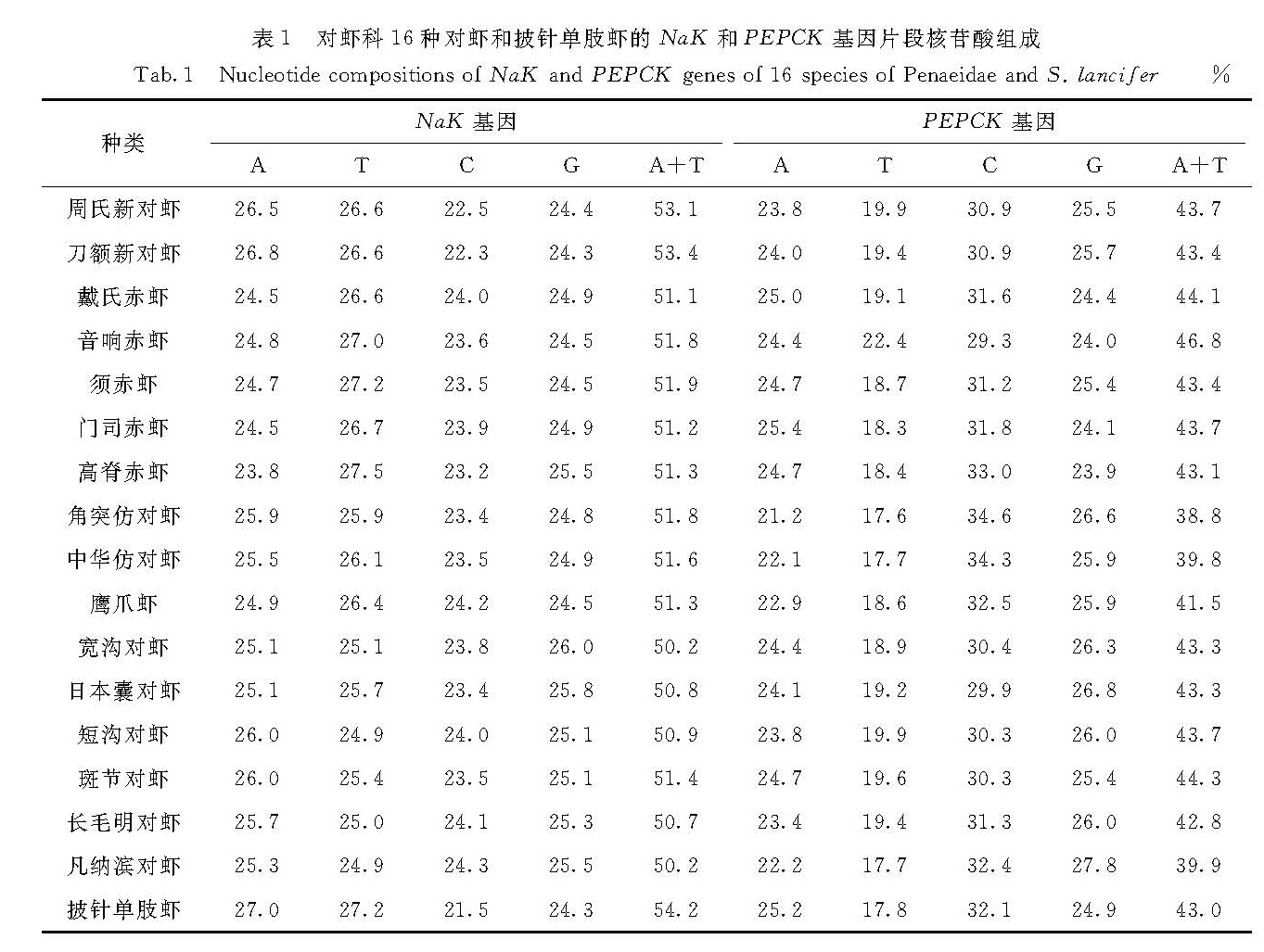
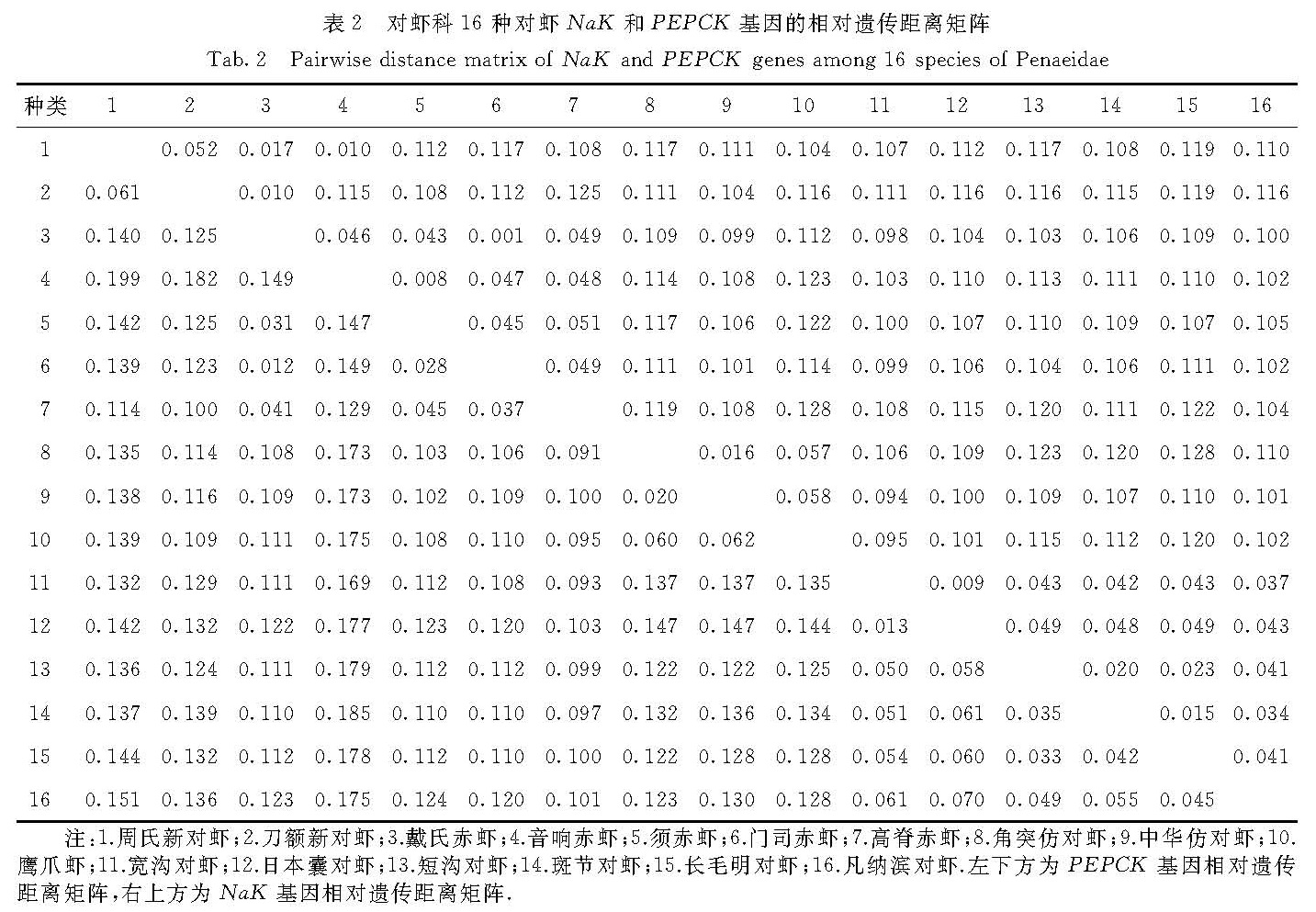
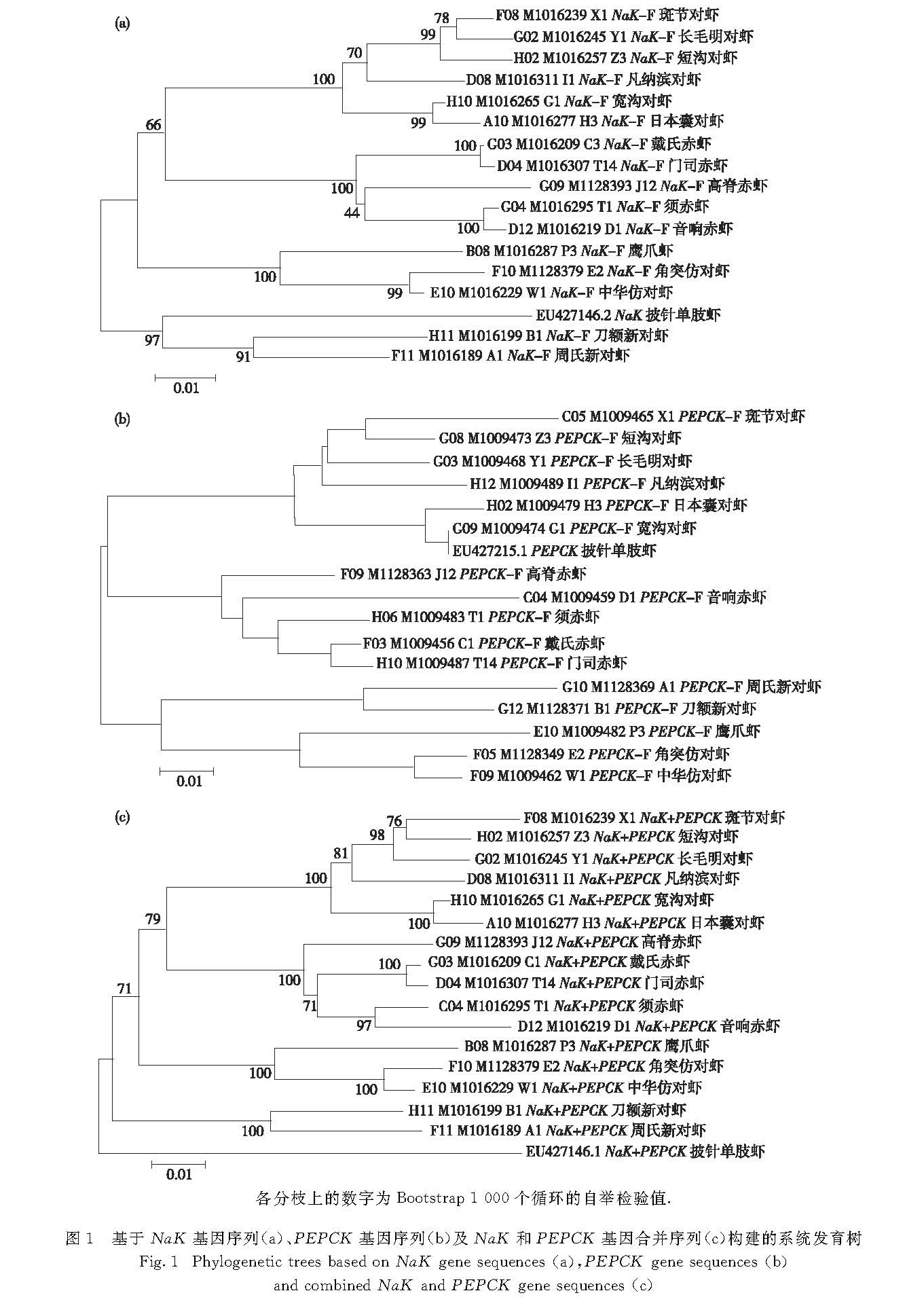
3.1 核基因的优势
本研究基于NaK基因序列建立的系统发育树中,对虾属的斑节对虾和短沟对虾会被长毛明对虾分开而不能聚为一枝; 但在基于PEPCK基因序列以及NaK和PEPCK基因合并序列构建的系统发育树中,对虾属的斑节对虾和短沟对虾能够聚为一枝.而之前的一些基于COI和16S rRNA基因序列的系统发育分析不能很好地解决斑节对虾与对虾属中其他对虾以及与明对虾属中对虾的关系,在所建的系统发育树中,对虾属的斑节对虾与短沟对虾总会被明对虾属的虾分开[11-12,20].因此,基于PEPCK基因序列以及NaK和PEPCK基因合并序列的分析能很好地解决这一问题,也很符合形态学上通过有无肝脊区分对虾属(有肝脊)和明对虾属(无肝脊)的结果[21].
赤虾属对虾的形态学主要分类特征是头胸甲后方有无下缘响器[21].本研究中赤虾属的音响赤虾和须赤虾头胸甲后缘具有响器,高脊赤虾、门司赤虾以及戴氏赤虾的头胸甲后缘无响器.在单独基于NaK基因和PEPCK基因序列构建的系统发育树中,高脊赤虾未能和具有相同形态特征的门司赤虾和戴氏赤虾聚在一起; 而基于PEPCK和NaK基因合并序列所构建的系统发育树中,音响赤虾和须赤虾聚为一枝,高脊赤虾、门司赤虾和戴氏赤虾则聚为另外一枝.因此,基于NaK和PEPCK基因合并序列的分析结果与形态学分类结果一致,并且其中门司赤虾和戴氏赤虾的关系相较于高脊赤虾更为亲近,这也符合形态学上高脊赤虾的额角及额角后脊呈鸡冠状而门司赤虾和戴氏赤虾的额角宽、下缘突出呈薄片状的特征[21].
综上所述,基于NaK和PEPCK基因合并序列的系统发育树在研究对虾的系统发育关系上有一定的优势.
3.2 对虾科9属对虾的系统发育关系
各分枝上的数字为Bootstrap 1 000个循环的自举检验值.
图1 基于NaK基因序列(a)、PEPCK基因序列(b)及NaK和PEPCK基因合并序列(c)构建的系统发育树
Fig.1 Phylogenetic trees based on NaK gene sequences(a),PEPCK gene sequences(b)
and combined NaK and PEPCK gene sequences(c)本研究选取了新对虾属、赤虾属、鹰爪虾属、仿对虾属、囊对虾属、沟对虾属、滨对虾属、明对虾属和对虾属的9属16种对虾,基于NaK和 PEPCK基因合并序列构建了系统发育树,由结果可知9个属的对虾除新对虾属外可以分为3个进化枝,这符合Burkenroad
[8]所提出的对虾科可以分成Penaeini、Trachypenaeini和Parapenaeini 3个群体的结果.1934年Burkenroad根据头胸甲额胃脊的有无以及额角侧沟的长短将对虾属(原)分为2个群体A和B
[22].1949年Kubo将群体B再划分为2个群体C(无肝脊)和D(有肝脊)
[7].1969年Farfante将这3个群体A、C、D分别命名为沟对虾亚属、明对虾亚属和滨对虾亚属,并且在滨对虾亚属中划分出对虾亚属
[23],1971年Tirmizi从沟对虾亚属中建立了日本囊对虾亚属
[24],1972年Burukovsky从美国的沟对虾亚属中建立了美对虾亚属,1997年Farfante和Kensley将广义的对虾属下的6个亚属提升为属
[2].
本研究中基于NaK和PEPCK基因合并序列构建的系统发育树的第一进化枝包含对虾属、明对虾属、滨对虾属、沟对虾属和囊对虾属,属于Penaeini群体,其中囊对虾属和沟对虾属聚为一枝,表明二者亲缘关系接近,这与形态学分类结果一致; 滨对虾属、明对虾属和对虾属聚为另外一枝,但对虾属在分子水平上相较于滨对虾属而言,与明对虾亲缘关系更近,这与形态学分类结果有差异.分子系统发育研究表明当前的形态学分类可能不能很好地显示对虾属、明对虾属和滨对虾属之间的亲缘关系,有待进一步研究.第二进化枝包含赤虾属,属于Parapenaeini群体.第三进化枝包含鹰爪虾属和仿对虾属,属于Trachypenaeini群体,而新对虾属在第三进化枝外.Voloch等[11]使用16S rRNA和COI基因研究对虾的系统发育关系中新对虾属也未和鹰爪虾属、仿对虾属聚在一起,因此,对于新对虾属的分类地位还应进行更多的研究.
致谢:非常感谢厦门大学海洋与地球学院的王德祥老师在形态学鉴别对虾过程中给予的帮助.