收稿日期:2016-12-15 录用日期:2017-05-15
基金项目:厦门市科技计划项目(3502Z20153005); 厦门南方海洋研究中心项目(15GYY024NF03)
通信作者:xpling@xmu.edu.cn
基金项目:厦门市科技计划项目(3502Z20153005); 厦门南方海洋研究中心项目(15GYY024NF03)
通信作者:xpling@xmu.edu.cn
(厦门大学化学化工学院,福建 厦门 361005)
(College of Chemistry and Chemical Engineering,Xiamen University,Xiamen 361005,China)
Mortierella alpina; arachidonic acid(ARA); baffled flask; B-group vitamins
DOI: 10.6043/j.issn.0438-0479.201612022
以本课题组自主分离并鉴定的一株花生四烯酸(arachidonic acid,ARA)生产菌株,即高山被孢霉(Mortierella alpina)LU166为研究对象,首先利用凹槽摇瓶培养,较为稳定地将菌体形态控制为刺突状小球,这一形态比絮状和大球状更有利于油脂和ARA的积累.在此基础上优化了4种B族维生素(VB1、VB12、生物素和泛酸钙)的添加量,并复合添加优化后的维生素浓度,最终获得的菌体生物量、油脂和ARA产量分别达到24.74,12.66和6.46 g/L,相比对照组分别提高了10%,30%和16%.
An arachidonic acid(ARA)-producing strain Mortierella alpina LU166,which is independently isolated by our lab,was studied in the present work.Firstly,it's found that the mycelial morphology can be steadily kept as fluffy small pellets when cultured in baffled flask,which is more favorable for the accumulation of total lipids and ARA than that by filaments and big pellets morpho-logy.Then,the concentration of B-group vitamins(VB1,VB12,biotin and calcium pantothenate)was optimized to enhance the lipid production.When adding the combination of vitamins with the optimized concentration into the culture,the biomass,lipids and ARA yield reached 24.74,12.66 and 6.46 g/L,respectively,which increased by 10%,30% and 16%,compared to the control group.
花生四烯酸(arachidonic acid,ARA)即全顺式二十碳-5,8,11,14-四烯酸,是一种ω-6高级不饱和脂肪酸,具有促进婴幼儿脑部发育、调节神经传导、降低胆固醇等多种重要的生理功能,在食品、医药、化妆品等领域有广泛应用[1].传统的ARA主要来源于动物油等[2],难以满足市场需求,利用微生物发酵法生产ARA具有周期短、不受地域限制等优点,成为当前研究的热点之一,而高山被孢霉(Mortierella alpina)是ARA的优良生产菌株[3].
丝状真菌在深层发酵过程中存在形态难以控制的难题.受环境和生长条件的影响,高山被孢霉会形成不同的菌体形态,如絮状、光滑小球、刺突状小球、松散羽状,或者成团.而菌体形态又极大地影响着发酵中的传质和溶氧(dissolved oxygen,DO)等过程,导致高山被孢霉产油性能的差异[4-7].因此,控制良好的菌体形态对于高山被孢霉发酵产ARA有重要意义.Park等[8]在高山被孢霉的形态学方面进行了相关研究工作,发现培养环境如温度、DO等对菌丝生长、菌球成核有极大的影响.已有研究[4-7]表明:菌体为中空光滑的小球形态时不利于其生长; 菌体为丝状时会增加培养基的黏度,阻碍传质; 刺突状小球为较佳的菌体生产形态,当菌球直径为1~2 nm时其ARA产量最高.高山被孢霉经脂肪酸合成(fatty acid synthesis,FAS)途径合成多不饱和脂肪酸,途径中脂肪酸脱饱和是一个耗氧过程,因此高山被孢霉产ARA需要一定的高DO环境,而菌球形态随DO的增加不断改变.形态学研究[9]结果表明:当环境中DO为10~15 mg/L时,培养基中菌体主要为直径小于1.5 mm的菌球和分散菌丝体; DO为20~30 mg/L时,菌体呈1.5~3.5 mm的蓬松状小球; 而过高的DO(40~50 mg/L)会导致菌球刺突溶解,形成中空光滑小球.
目前,对高山被孢霉发酵生产ARA的基础培养条件的研究较多,主要集中在碳源、氮源的优化[10-11],钙、镁等矿物质对高山被孢霉发酵生产ARA的影响[6],而维生素对其的影响研究相对较少.任路静等[12]研究发现生物素(VB7)作为乙酰辅酶A(乙酰-CoA)羧化酶的羧基载体,可以促进裂殖壶菌(Schizochytrium sp.)油脂积累.作为产油微生物,高山被孢霉发酵合成ARA的过程可分为细胞生长阶段和油脂合成阶段[13],在油脂合成阶段,长链不饱和脂肪酸由乙酰-CoA经FAS途径合成,因此,乙酰-CoA是油脂合成的重要前体.泛酸钙(VB5)是CoA的组成成分,维生素B12(VB12)参与催化L-甲基丙二酸单酰-CoA[14]; VB7在酶促羧化反应中作为活动羧基载体,是以碳酸氢盐作为羧化剂的羧化反应中的必要辅酶,参与乙酰-CoA羰化为丙二酰-CoA等过程[15].可见多种B族维生素是ARA合成过程的重要因子.
因此,本文中研究了凹槽摇瓶和普通平底摇瓶培养对高山被孢霉的形态调控和合成ARA的影响.通过初筛确定了4种对高山被孢霉产ARA有促进作用的B族维生素(VB1、VB12、VB7、VB5),并对这4种维生素的添加浓度进行优化,考察其对高山被孢霉菌体生长、油脂和ARA积累的影响,以期提高高山被孢霉合成ARA的代谢通量.
高山被孢霉菌株LU166,由厦门大学生物化工研究所及中国微生物菌种保藏中心保藏(CGMCC No.12764).
PDA固体培养基(g/L):马铃薯200,葡萄糖20,琼脂20.种子培养基(g/L):葡萄糖30,酵母粉4,KH2PO4 2,MgSO4·7H2O 0.3,pH 6.0.发酵培养基(g/L):葡萄糖50,酵母粉8,玉米浆3,谷氨酸1,KH2PO4 2,MgSO4·7H2O 0.1,ZnSO4·7H2O 0.1,CaCO3 0.05,1 000×微量元素1 mL,pH 6.0.其中,1 000×微量元素组成(g/L):MnCl2·4H2O 0.8,H3BO30.5,FeCl3·6H2O 0.2,NiSO4·6H2O 0.05,CoCl2·6H2O 0.005,CuSO4·5H2O 0.002,置于4 ℃下保存.
凹槽摇瓶由250 mL平底摇瓶经热加工为瓶底向内凹陷形成4个1 cm左右的“玻璃挡板”,形状如图1所示.

图1 凹槽摇瓶(a)与平底摇瓶(b)
Fig.1 The shapes of baffled flask(a)and normal flask(b)
从实验室保存的菌种中分离出2株形态类似于高山被孢霉的菌株(菌1和菌2).采用转录间隔区(ITS)设计引物(ITS-F:5'-GAAGGATCATTCATAATCAA-3',ITS-R:5'-GATTTGAGATCGAGTTTCAA-3')扩增,其中菌1用ITS引物的扩增结果为阴性,故采用18S rRNA基因通用引物扩增.
取5 mL发酵液,用布氏漏斗真空抽滤,分离发酵液和菌体,菌体用去离子水洗涤3次后,放置于15 mL离心管中于-20 ℃下冷冻干燥,12 h后取出称量.菌体生物量计算公式如下:
ρB=(m-m0)/V,
其中,ρB为菌体生物量,m为冷冻干燥后总质量,m0为空管质量,V为发酵液取样体积.
将冷冻干燥后的菌体放入装有磁子的离心管中,加入4 mL 38%(质量分数)盐酸,用恒温金属浴锅(HTPOT 10)于65 ℃水浴下搅拌破碎1 h.再加入5 mL正己烷,摇匀萃取5 min,静置分层,取上清液至离心管中,重复萃取至上清液为无色.将上清液用氮气吹扫干燥至恒量(m1).油脂产量计算公式如下:
ρTL=(m1-m2)/V,
其中,ρTL为油脂产量,m2为空管质量.
向提取的油脂样品中加入5 mL 0.5 mol/L KOH-甲醇溶液,65 ℃水浴直至油滴消失.加入5 mL 30%(质量分数)三氟化硼乙醚,65 ℃水浴30 min,取出冷却.精确加入5 mL正己烷和50 μL十七烷酸甲酯,再加入1 mL饱和NaCl溶液,震荡萃取15 min,静置分层.取上清液,经无水硫酸钠除水,待测.
气相色谱检测条件:SP-2560气相毛细管柱(100 m×0.25 mm×0.2 μm),初始柱温为140 ℃,检测器温度为260 ℃,柱箱温度为140 ℃,载气为氮气.升温程序:初始温度140 ℃,维持5 min,再以4 ℃/min速率升温至240 ℃,维持30 min.样品中ARA的质量浓度根据下式计算:
ρARA=SARA×ρ17×FARA/S17,
其中,ρARA和ρ17分别表示ARA和内标十七烷酸甲酯的质量浓度,SARA和S17表示气相谱图中ARA和十七烷酸甲酯的峰面积,FARA表示ARA标准品相对于内标的响应因子.
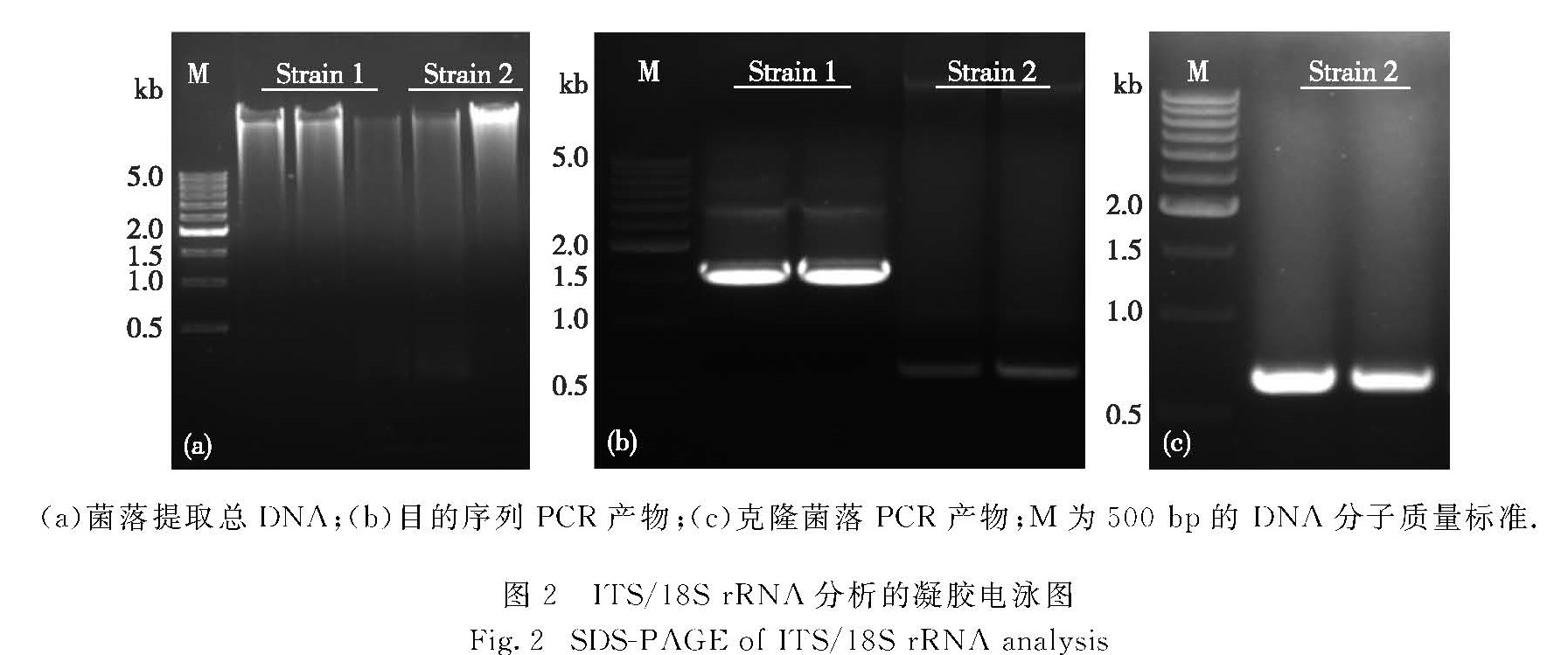
图2 ITS/18S rRNA分析的凝胶电泳图
Fig.2 SDS-PAGE of ITS/18S rRNA analysis

图3 高山被孢霉LU166的ITS序列
Fig.3 ITS sequence of M. alpina LU166
分别用试剂盒提取两株菌的总DNA(图2(a)),经PCR,菌1得到约1 500 bp的18S rRNA基因扩增片段,菌2得到约600 bp的ITS扩增片段(图2(b)).割胶
回收,经T载体连接转化后涂板(含氨苄青霉素),菌2得到白色克隆,菌1结果为阴性.挑取菌2的克隆菌落进行菌落PCR鉴定,结果为阳性(图2(c)).对菌1总DNA的PCR扩增片段割胶回收产物以及菌2的克隆菌落培养液进行测序.
将上述测序所得序列提交至GenBank进行Blast同源性检索.结果表明:菌1与高山被孢霉菌不匹配; 菌2与多株高山被孢霉菌有99%相似性,故鉴定菌2为高山被孢霉,并命名为高山被孢霉LU166,其ITS序列如图3所示.利用N-J方法构建系统进化树见图4,结果表明高山被孢霉LU166与CBS 210.32(JN943803)进化关系最为密切.
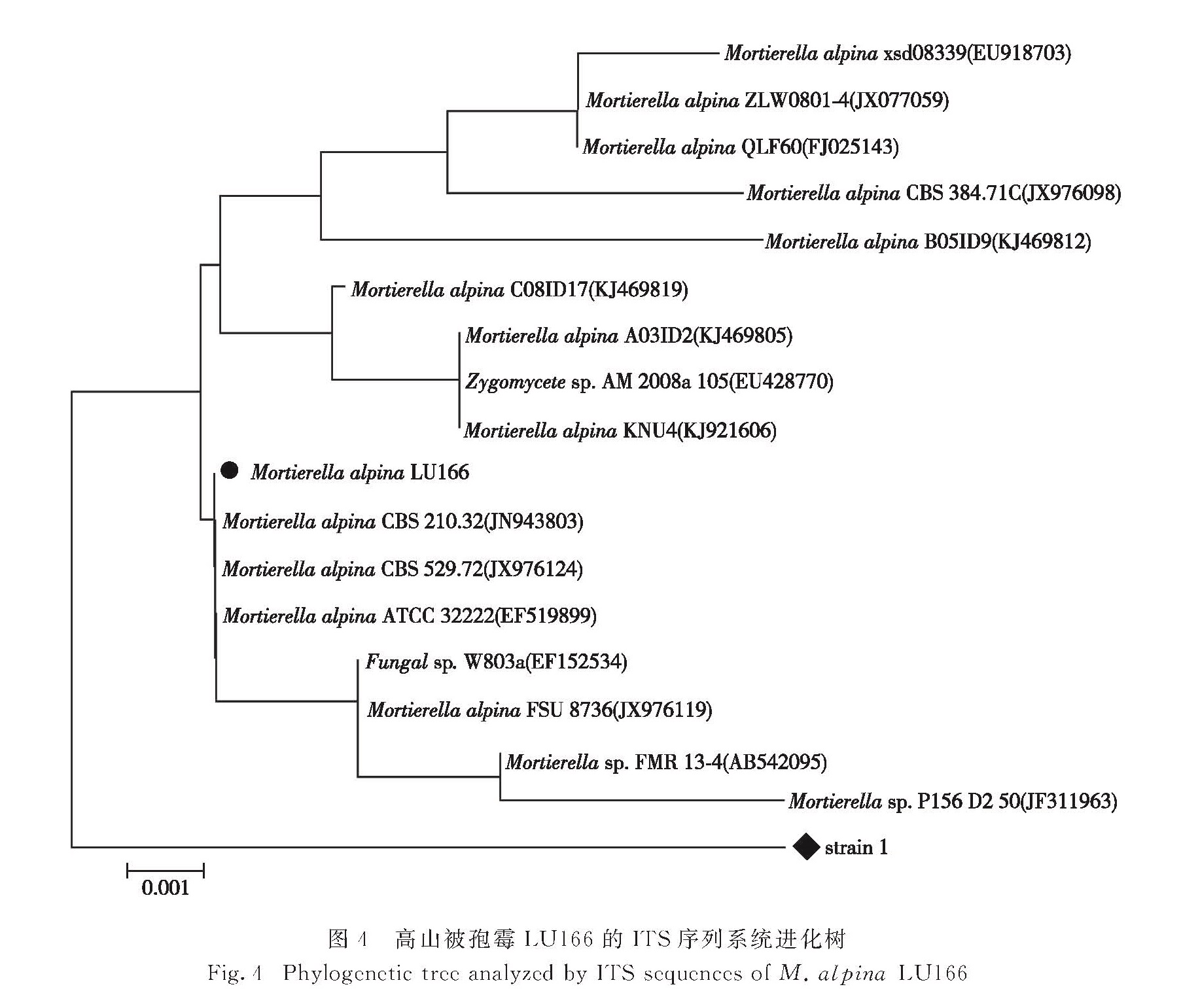
图4 高山被孢霉LU166的ITS序列系统进化树
Fig.4 Phylogenetic tree analyzed by ITS sequences of M. alpina LU166
在不同摇瓶中培养高山被孢霉LU166,观察其形态,结果如图5所示.在其他条件相同时,采用平底摇瓶和凹槽摇瓶培养下,菌体呈现出不同的形态.多次摇瓶发酵后发现,采用平底摇瓶培养获得的菌体形态不稳定:有时为分散的自由菌丝体和絮状体(图5(a)),有时形成直径为4~5 mm的刺突状大球(图5(b)),有的甚至无法分散而聚集成团(图5(c)).而凹槽摇瓶培养下,菌体普遍为2 mm左右带刺突的小球(图5(d)),形态相对较为稳定.
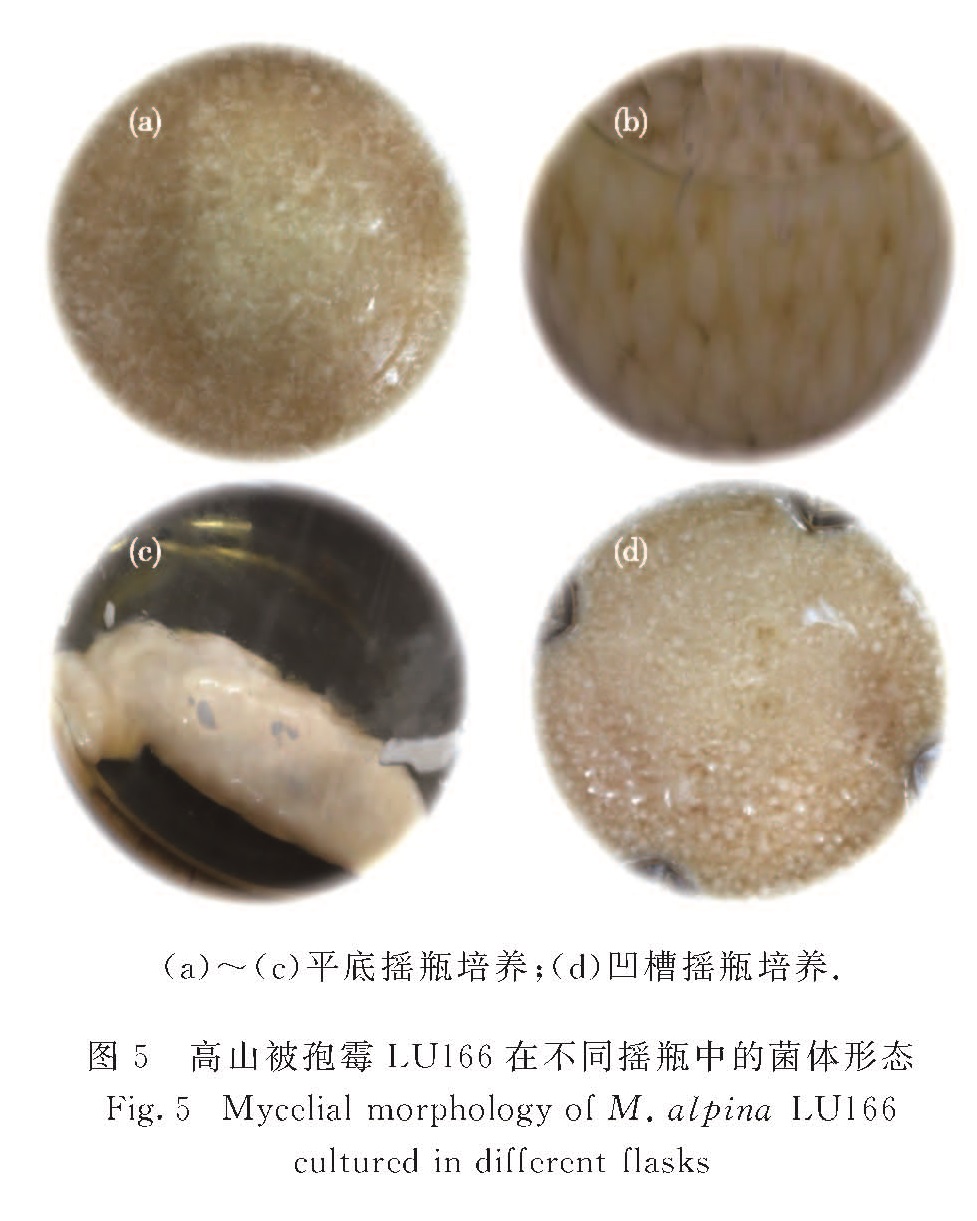
图5 高山被孢霉LU166在不同摇瓶中的菌体形态
Fig.5 Mycelial morphology of M. alpina LU166 cultured in different flasks
高山被孢霉LU166在不同形态下的菌体生物量、油脂和ARA的产量及油脂成分如图6所示.可以看出,菌体形态不同对高山被孢霉菌体生长、油脂和ARA积累情况的影响有很大的差异.大球状、絮状及刺突状小球形态下,菌体生物量相差不大,油脂产量也没有明显的差别,但ARA产量则有明显差异(图6(a)).刺突状小球形态下ARA在生物量和油脂中的比例明显高于大球状和絮状形态下的结果(图6(b)).其中,刺突状小球菌体的ARA产量和含量均比大球状菌体提高了102%.3种形态下油脂成分(图6(c))的变化主要体现在饱和脂肪酸(C16:0和C18:0)、油酸(C18:1)和ARA的含量变化,而其他油脂成分的含量变化不大,这说明ARA的增加是由于饱和脂肪酸和油酸在代谢途径中被有效地转化为ARA.结合图5的菌体形态分析,凹槽摇瓶向内凹陷的“玻璃挡板”给菌体提供了一定的冲击力,使得DO增加,从而有利于菌丝生长和尖端形成,并使菌体分散而不易形成严重的聚团现象,且这种“玻璃挡板”不同于发酵罐叶片,不会造成大的剪切力,菌体保持较为完整的刺突状小球,该形态有1和2为采用平底摇瓶培养获得的大球状和絮状菌体; 3为采用凹槽摇瓶培养获得的刺突状小球菌体.

图6 不同摇瓶对高山被孢霉LU166发酵的影响
Fig.6 Effects of different flasks on the fermentation of M. alpina LU166
利于高山被孢霉产ARA[4-7].平底摇瓶相较于凹槽摇瓶的DO较低,当DO低于菌丝体生长最适值时,菌丝生长速率、分支尖端形成和尖端延伸速率都降低,这种情况下菌体多产生短的自由菌丝体和絮状体,而絮状形态下菌液黏度较大,进一步增大了传质阻力,不利于氧传递; 大球状菌体因为菌球太大,虽然菌球表面传质良好,但越靠近球
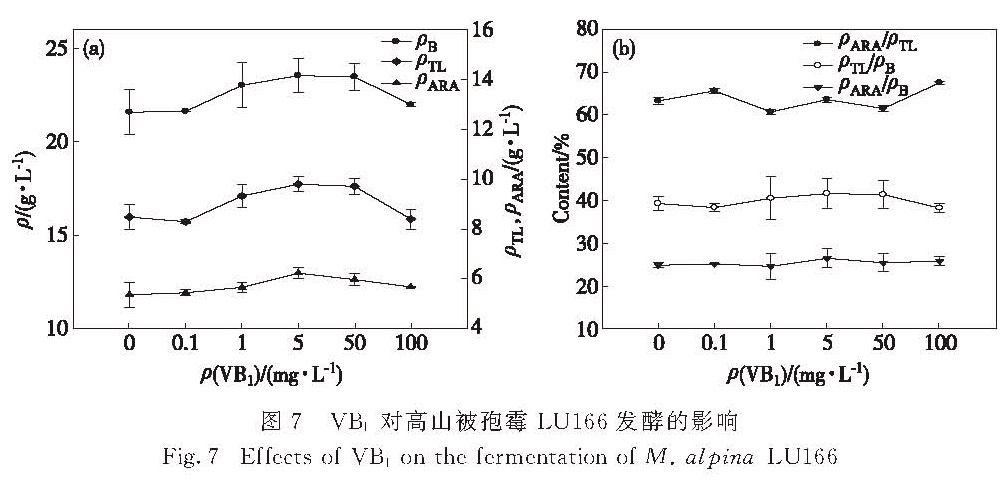
图7 VB1对高山被孢霉LU166发酵的影响
Fig.7 Effects of VB1 on the fermentation of M. alpina LU166
核中心,基质和氧气越少,球心处甚至会出现基质和氧气耗尽的情况.FAS途径中合成不饱和脂肪酸的去饱和作用所需
的DO要高于菌体生长和油脂积累的需氧量,因此在平底摇瓶条件下,偏低的DO没有抑制菌体生长和油脂积累,但极大地降低了ARA在总脂肪酸中的比例.特别是在大球状形态下,由于菌核内部缺氧,ARA的合成被明显抑制,这与之前的报道观点[15]相一致.综合来看,刺突状小球是高山被孢霉产ARA的最佳形态,因此,选择凹槽摇瓶进行后续实验.
在发酵培养基中分别添加0.1,1,5,50,100 mg/L的VB1,考察其对高山被孢霉LU166的菌体生物量、油脂和ARA产量的影响,结果如图7所示.在培养基中添加VB1,促进了菌体的生长,提高了油脂和ARA的产量,当VB1为5 mg/L时,生物量、油脂和ARA产量最高,分别比对照组提高了9%,15%和11%,随着VB1质量浓度继续增加,相关产量开始下降(图7(a)).另外,VB1的添加对ARA在油脂中的比例影响不大,但在一定程度上提高了油脂和ARA占菌体生物量的比例,且添加5 mg/L VB1的提高效果最明显(图7(b)).这说明添加适宜质量浓度的VB1可以促进菌体合成油脂.
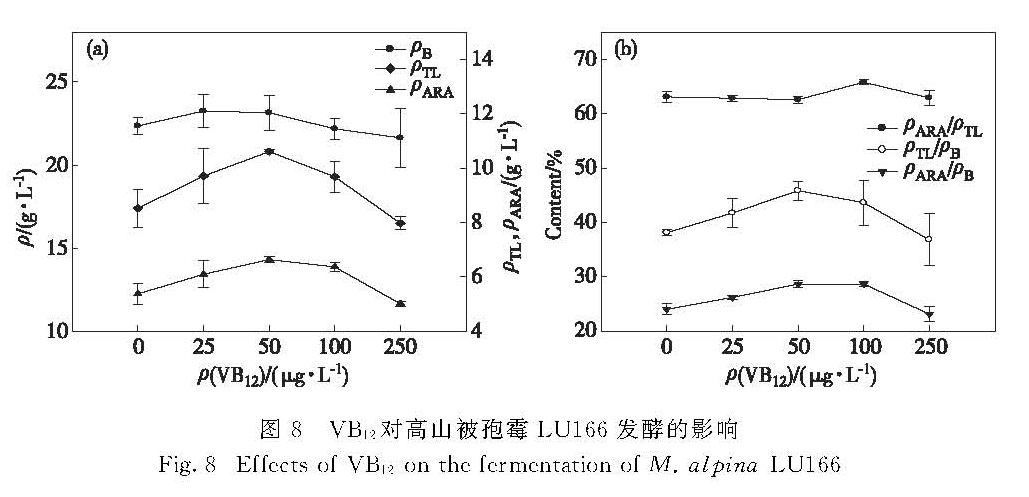
图8 VB12对高山被孢霉LU166发酵的影响
Fig.8 Effects of VB12 on the fermentation of M. alpina LU166
在发酵培养基中分别添加25,50,100和250 μg/L的VB12,考察其对高山被孢霉LU166的菌体生物量、油脂和ARA产量的影响,结果如图8所示.由图8(a)可知:VB12的添加对菌体生长的促进作用不明显,但对油脂和ARA的积累有明显的促进作用; 当VB12质量浓度为50 μg/L时,油脂和ARA的产量达到最大值,分别比对照组提高了24%和23%; 当继续增加VB12的质量浓度时,菌体生长、油脂和ARA的积累受到抑制.图8(b)的结果表明VB12的添加提高了油脂和ARA占菌体生物量的比例,在VB12质量浓度为100 μg/L时,ARA占油脂的比例有所增加,这是由于此时油脂的产量比较低而导致的.
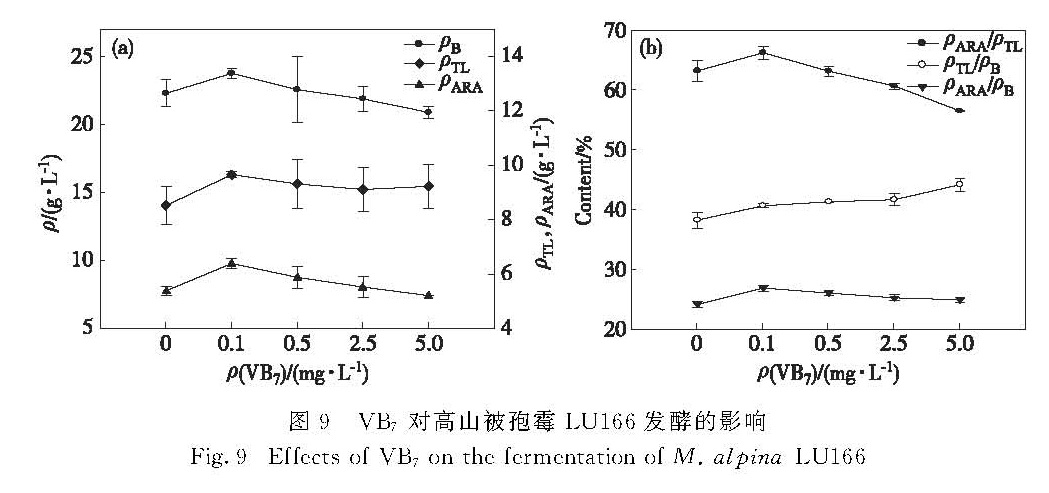
图9 VB7对高山被孢霉LU166发酵的影响
Fig.9 Effects of VB7 on the fermentation of M. alpina LU166
在发酵培养基中分别添加0.1,0.5,2.5和5.0 mg/L的VB7,考察其对高山被孢霉LU166的菌体生物量、油脂和ARA产量的影响,结果如图9所示.添加低质量浓度的VB7,对菌体生长、油脂和ARA的积累有促进作用,并在一定程度上提高了油脂和ARA占菌体生物量的比例,且VB7的添加对油脂占菌体生物量比例的提升高于ARA占菌体生物量的比例,因此ARA占油脂比例呈现出下降趋势.VB7的质量浓度为0.1 mg/L时促进作用最佳,菌体生物量、油脂和ARA产量分别比对照组提高了7%,13%和19%.
在发酵培养基中分别添加0.5,1.25,2.5,5.0和10 mg/L的VB5,考察其对高山被孢霉LU166的菌体生物量、油脂和ARA产量的影响,结果如图 10所示.在培养基中添加VB5后,明显促进了菌体生长,但对油脂和ARA的积累的促进效果不明显,因此油脂和ARA占菌体生物量的比例呈下降趋势.VB5为0.5~2.5 mg/L时抑制ARA的积累; VB5为5.0 mg/L时对菌体生长的促进作用最为明显,菌体生物量比对照组提高了20%,油脂和ARA的产量则分别比对照组提高了7%和6%,可见,此时油脂和ARA产量的提高归因于菌体生物量的增大; 继续增加VB5质量浓度至10 mg/L时则对菌体生长、油脂和ARA的积累均产生抑制作用.
将4种维生素在各自最佳条件下的发酵数据统计在表1中,并通过最小显著性差异法(LSD)对发酵结果进行显著性分析.可以看出,不同维生素对菌体生物量、油脂和ARA产量的影响有所差异.其中,VB12对油脂和ARA的积累的促进作用最明显,VB12的质量浓度为50 μg/L时ARA的产量与对照组相比具有显著性差异,VB1和VB7对ARA的产量有一定的提升效果,而VB5对菌体生长的促进作用较好.
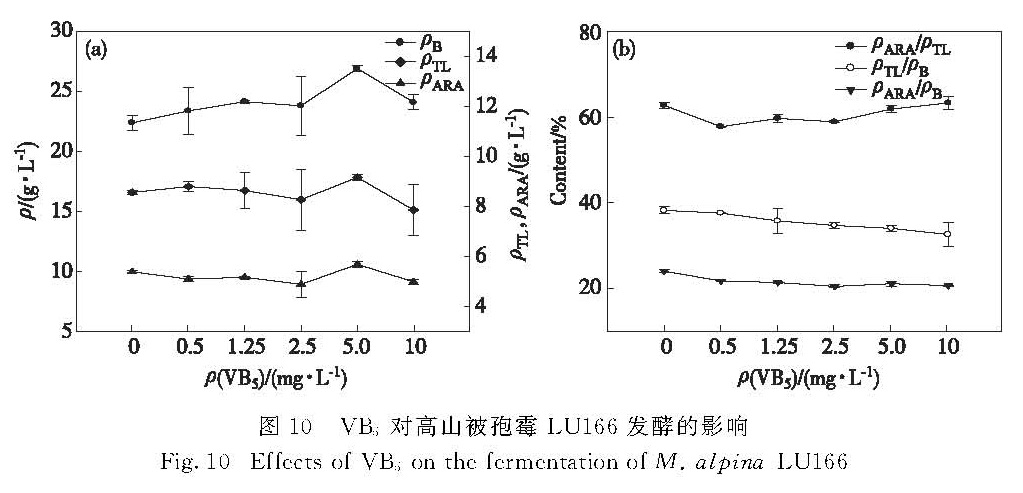
图 10 VB5对高山被孢霉LU166发酵的影响
Fig.10 Effects of VB5 on the fermentation of M. alpina LU166
根据以上分析结果,实验进一步复合这4种维生素的最优浓度进行高山被孢霉LU166的发酵,结果如图 11所示.复合维生素对菌体的生长代谢有明显的促进作用,最终菌体生物量、油脂和ARA产量分别达到24.74,12.66和6.46 g/L,分别比对照组提高了10%,
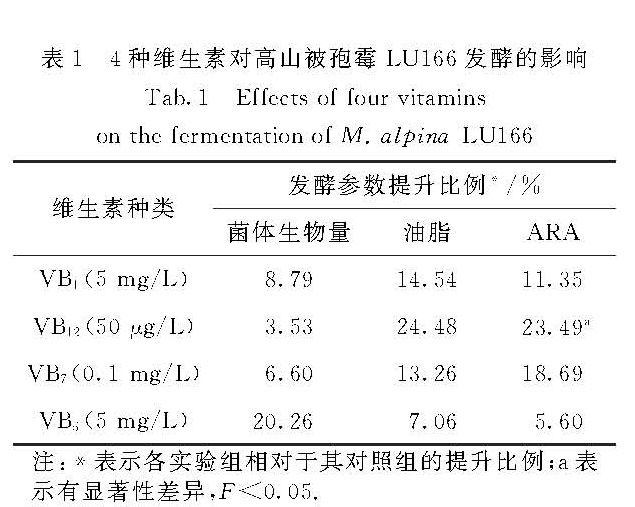
表1 4种维生素对高山被孢霉LU166发酵的影响
Tab.1 Effccts of four vitamins on the formcntation of M.alpina LU166
30%和16%; 油脂和ARA占菌体生物量的比例分别达到51%和26%,分别比对照组提高了18%和6%.与单因素维生素添加实验相比,复合维生素的添加使油脂和ARA产量进一步提高.同时,ARA占油脂的比例有所下降,这可能是因为添加的这4种B族维生素更多地参与脂肪酸合成的前期过程,对脱饱和作用没有明显的促进作用.

图 11 复合维生素对高山被孢霉LU166发酵的影响
Fig.11 Effects of the mixed vitamins on the fermentation of M. alpina LU166
本研究利用凹槽摇瓶培养形态难以控制的高山被孢霉菌株发酵生产ARA,凹槽挡板带来的冲击有利于高山被孢霉稳定形成刺突状小球形态,这一形态有利于高山被孢霉产ARA[4-7]; 同时,凹槽挡板也为FAS途径中脂肪酸去饱和酶催化脱氢提供较高的DO,提高了油脂转化为ARA的能力.与平底摇瓶培养下形成的大球状菌体相比,凹槽摇瓶培养下形成的刺突状小球菌体在ARA的产量和含量均提高了102%.在此基础上,添加适宜浓度的B族维生素VB1、VB12、VB7和VB5对高山被孢霉LU166的菌体生长、油脂和ARA的积累有一定的促进作用,优化后,复合B族维生素的添加使菌体生物量、油脂和ARA的产量分别达到24.74,12.66和6.46 g/L,分别比对照组提高了10%,30%和16%.该实验策略在发酵层面上提高了高山被孢霉产ARA的能力,对产油霉菌的发酵生产提供了一定的借鉴意义.今后的实验可以进一步结合基因工程手段,提高饱和脂肪酸对ARA的转化率,从而提高ARA的产量.
