2.1 乳酸对BG活力的影响
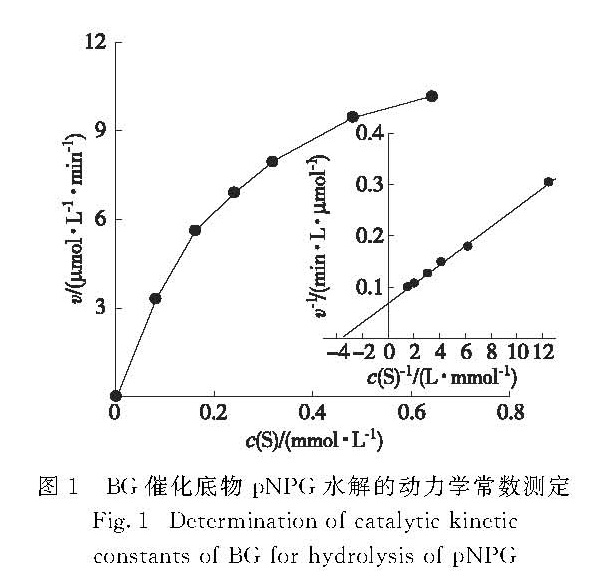
在酶的最适反应条件下(pH 5.0,60 ℃),首先测得东方肉座菌BG水解pNPG的米氏常数Km为0.28 mmol/L,最大反应速率Vm为2.1×104 U/mg(图1).
图1 BG催化底物pNPG水解的动力学常数测定
Fig.1 Determination of catalytic kinetic constants of BG for hydrolysis of pNPG
向测活体系中加入不同体积的乳酸,研究不同体积分数的乳酸对BG活力的影响.BG活力与乳酸体积分数的关系见图2:BG活力随着乳酸体积分数的增大而迅速下降,当乳酸体积分数为1.5%时,BG活力下降50%,即得乳酸对BG活力的半抑制浓度(IC50)为0.19 mol/L.
图2 乳酸对BG活力的影响
Fig.2 Effect of lactic acid on BG activity
2.2 乳酸对BG的抑制作用机制
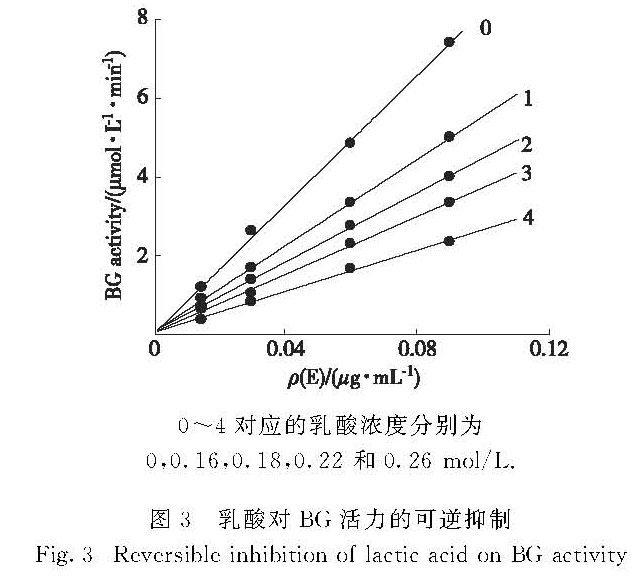
为判断乳酸对BG的抑制是否为可逆抑制,进一步在含不同浓度乳酸的测活体系中,固定底物pNPG浓度为0.24 mmol/L,改变BG的加入量,测定不同浓度乳酸对BG活力的影响.以酶活力对酶的加入量作图,得到一组通过原点的直线(图3),说明乳酸对BG的抑制作用是可逆过程.
图3 乳酸对BG活力的可逆抑制
Fig.3 Reversible inhibition of lactic acid on BG activity
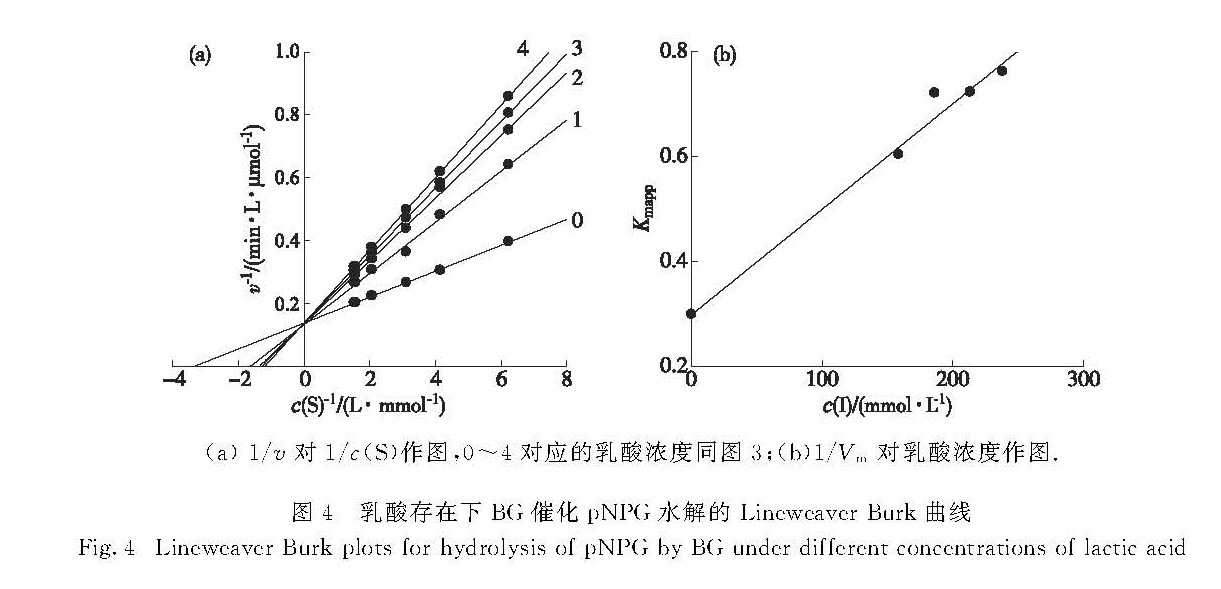
图4 乳酸存在下BG催化pNPG水解的Lineweaver-Burk曲线
Fig.4 Lineweaver-Burk plots for hydrolysis of pNPG by BG under different concentrations of lactic acid
在酶的最适反应条件下(pH 5.0,60 ℃),在含一定浓度的乳酸测活体系中改变底物浓度c(S),测定酶促反应的初始反应速率v,以1/v对1/c(S)进行双倒数作图,得到一组相交于纵轴的直线(图4(a)),说明乳酸不影响最大反应速率Vm,只影响米氏常数Km,其抑制类型表现为竞争性抑制,即乳酸与游离酶竞争性结合,但不与酶-底物复合物结合.以不同乳酸浓度下的表观米氏常数Kmapp对乳酸浓度作图得一直线(图4(b)),从斜率可求出其抑制常数KI为0.149 mol/L.
2.3 乳酸对BG的抑制动力学模型建立
参考文献[11-12]的方法建立乳酸对BG抑制动力学模型.根据实验结果,乳酸对酶作用为可逆竞争抑制,其动力学模型可表示为:
其中:E、S、I和P分别代表游离酶(BG)、底物(pNPG)、抑制剂(乳酸)和产物(pNP); ES为酶-底物复合物; EI为酶-抑制剂复合物; k+0与k-0分别表示酶的正、逆向速率常数; kp为酶-底物复合物分解速率常数.通常情况下,由于底物和抑制剂浓度远大于酶的初始浓度,根据邹承鲁教授提出的抑制剂对酶的抑制作用机制[13],可推导得下式:
c(P)t=(Bv)/(Ac(I)+B)·t+
(Ac(I)v)/((Ac(I)+B)2)(1-e-(Ac(I)+B)·t),(1)
其中,A=(Km·k+0)/(Vm+c(S)),(2)
B=k-0.(3)
式中,c(P)t是反应时间t时的产物浓度,A和B分别为抑制剂对酶的表观正向速率常数和逆向速率常数.当反应时间足够长,曲线变直,酶受抑制达到稳态,理论计算所得的产物浓度c(P)calc可表示为:
c(P)calc=(Bv)/(Ac(I)+B)·t+(Ac(I)v)/((Ac(I)+B)2),(4)
c(P)calc-c(P)t=(Ac(I)v)/((Ac(I)+B)2)·e-(Ac(I)+B)·t.(5)
分别对式(5)两边取自然对数,则得:
ln(c(P)calc-c(P)t)=a-(Ac(I)+B)·t,(6)
其中a为常数.
不同乳酸浓度下,以ln(c(P)calc-c(P)t)对反应时间t作图,可得一组直线,其负斜率(-slope)为Ac(I)+B,即斜率与抑制剂浓度呈线性关系,为一组交于纵轴的直线,其纵轴截距和斜率分别为B(即k-0)和A.联解式(2)和米氏方程,得到:
A/v=(Km·k+0)/(Vm)·1/(c(S)).(7)
以A/v对1/c(S)作图,直线的斜率为(Km·k+0)/Vm,由于Km与Vm已测出,所以从直线的斜率即可求得正向速率常数k+0.
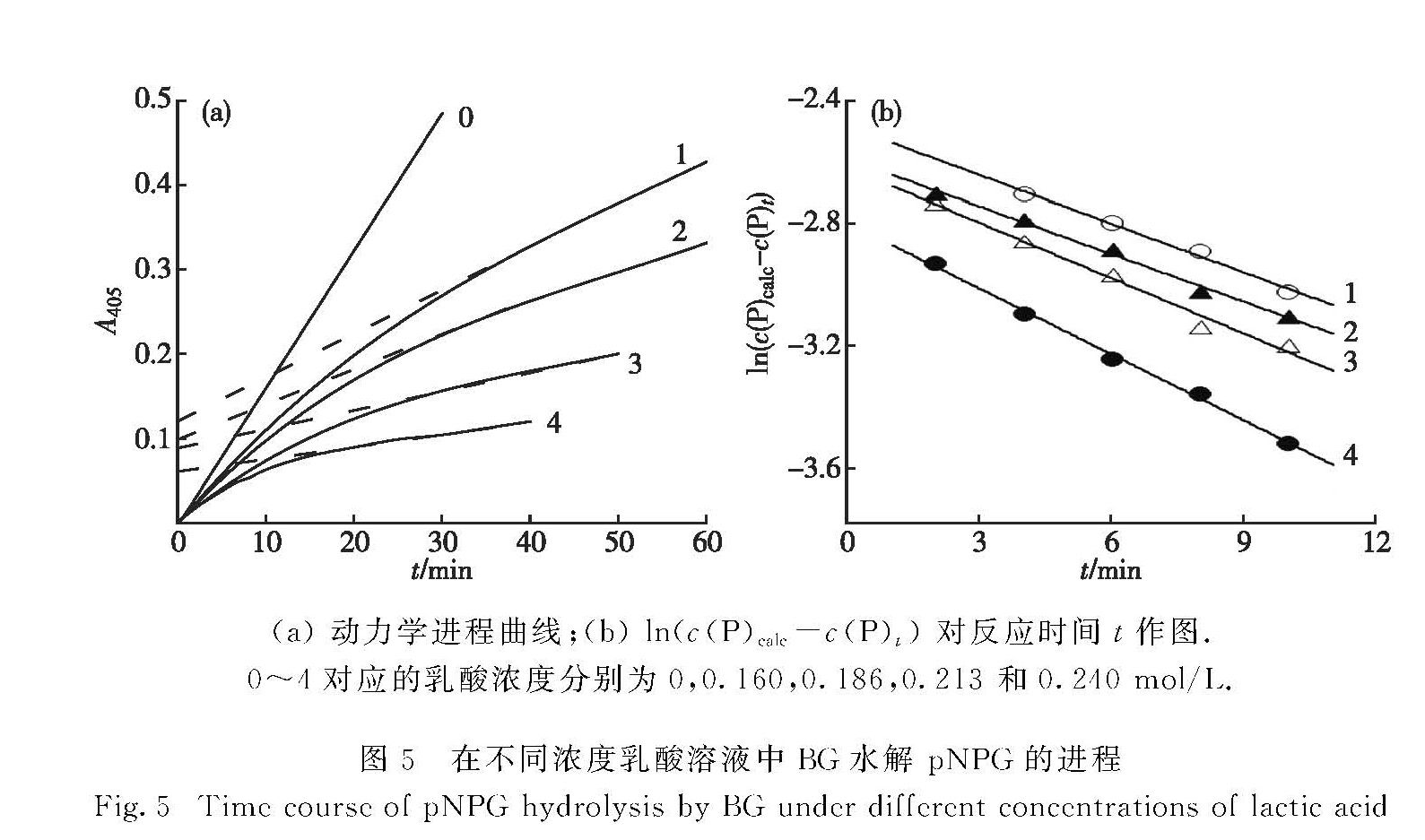
在pH 5.0的测活体系中(1 mL缓冲体系内含0.24 mmol/L pNPG及不同浓度的乳酸)加入适量酶液,在60 ℃下监测产物形成量(用A405表示)随反应时间t的进行过程.如图5(a)所示:在反应开始时,随着反应时间的增加,产物增加较为明显,随后产物增加逐渐缓慢直至达到恒定直线上升,说明酶逐渐被抑制,最终保留一定的剩余活力,且剩余活力随着乳酸浓度的增大而下降.可见,乳酸在考察浓度范围内对酶的效应表现为慢可逆的抑制作用.以ln(c(P)calc-c(P)t)对反应时间t作图,得一组直线(图5(b)),其负斜率即为Ac(I)+B.
图5 在不同浓度乳酸溶液中BG水解 pNPG的进程
Fig.5 Time course of pNPG hydrolysis by BG under different concentrations of lactic acid
2.4 乳酸对BG的微观速率常数测定
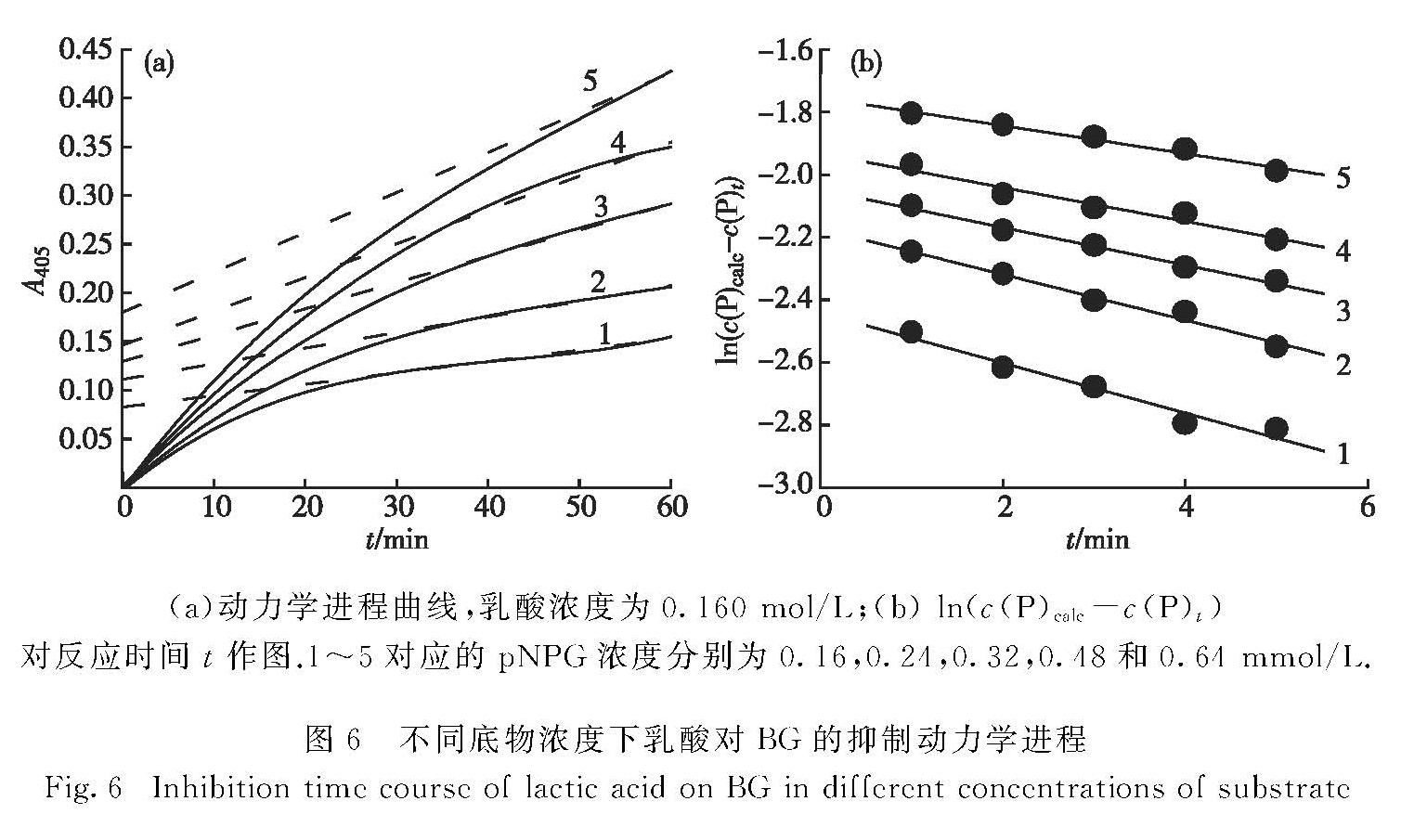
在含一定浓度乳酸的测活体系中,分别测定酶催化不同浓度pNPG(0.16,0.24,0.32,0.48和0.64 mmol/L)反应的动力学进程曲线,并以5个底物浓度反应曲线的ln(c(P)calc-c(P)t)对t作图,可得一组直线.图6(a)为酶在含0.160 mol/L乳酸的测活体系中不同底物浓度下催化pNPG水解的动力学进程曲线.以ln(c(P)calc-c(P)t)对t进行二次作图(图6(b)),得直线的负斜率为Ac(I)+B.
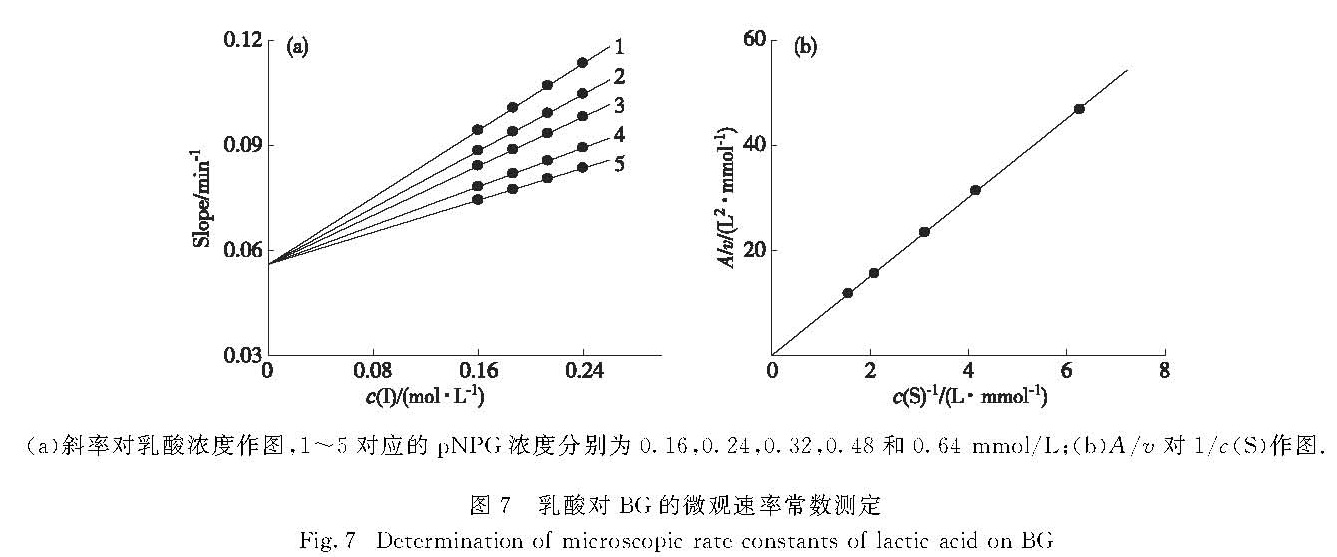
以直线斜率对乳酸浓度作图,可得到一组纵截距为B的直线(图7(a)),求出乳酸对BG的逆向速率常数k-0为0.056 min-1.根据已测定的不同底物浓度下的初始反应速率v及求得的A,以A/v对1/c(S)作图得到一条过原点的直线(图7(b)).根据式(7),由于Km、Vm及k-0已测出,所以从直线的斜率可求得乳酸对BG的正向微观抑制速度常数k+0为0.4 L/(mol·min).
图6 不同底物浓度下乳酸对BG的抑制动力学进程
Fig.6 Inhibition time course of lactic acid on BG in different concentrations of substrate