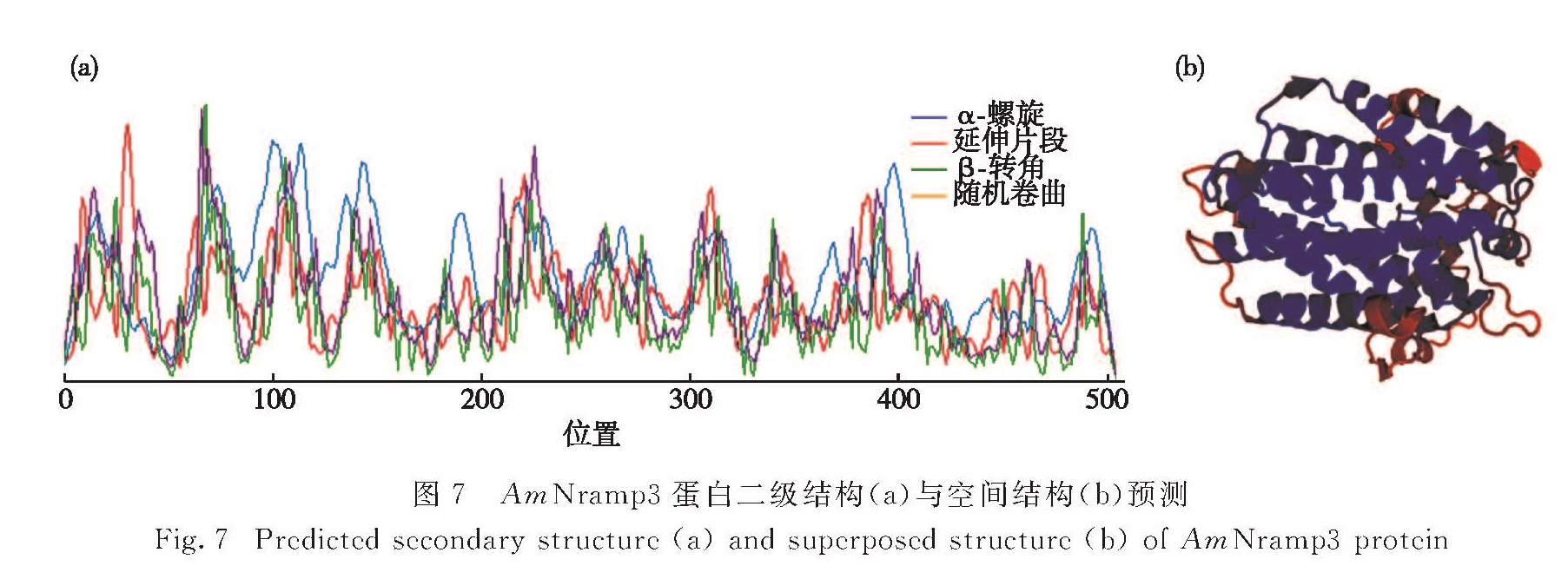
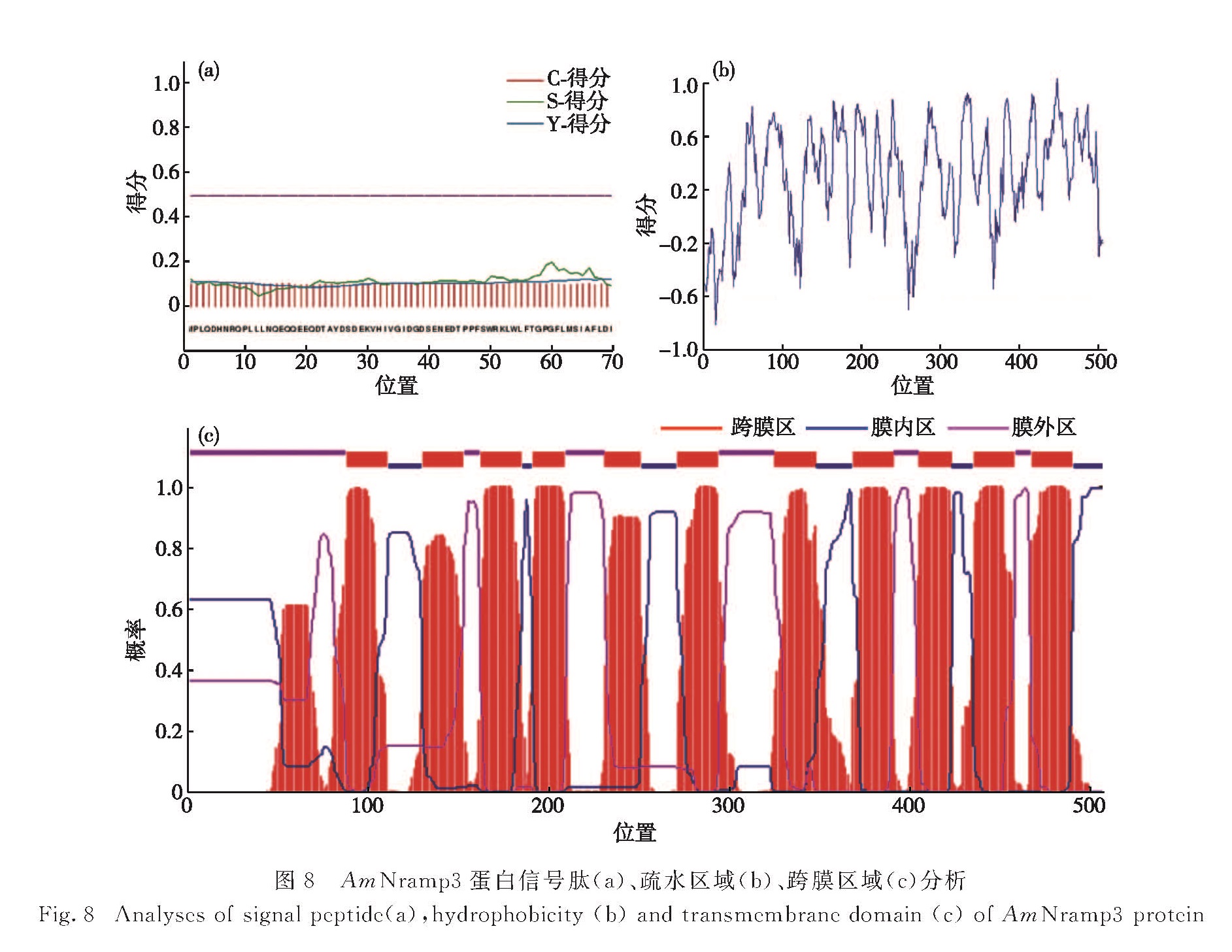
白骨壤(Avicennia marina(Forsk.)Vierh.)作为红树林的先锋植物,广泛分布于我国南海沿岸,是一种研究红树植物重金属污染物耐受性分子机制很好的材料.以抑制消减杂交获得的白骨壤重金属耐受性相关基因AmNramp3片段为基础,设计基因特异性引物,利用cDNA末端快速扩增(RACE)技术克隆了其全长,长度为1 958 bp; 通过实时荧光定量PCR(qRT-PCR)检测了AmNramp3在重金属镉胁迫时白骨壤叶、茎和根中的表达情况,其表达量在叶中高于根和茎中,但在茎和根中有快速的响应.利用生物信息学方法预测并分析了该基因的开放阅读框,位置位于1~1 527 bp,编码508个氨基酸; 进而预测并分析了AmNramp3蛋白的结构,发现其氨基酸序列中不含有信号肽,说明其不是分泌蛋白; 但其具有很复杂的跨膜结构,是一种跨膜蛋白.综上,AmNramp3基因编码一种跨膜蛋白,在重金属镉胁迫下表达量快速变化,可能与镉的跨膜运输密切相关,这为后续分析该基因发挥功能的分子机制奠定了基础.
The halophytic Avicennia marina(Forsk.)Vierh. is one of the pioneer mangroves along the southern coast of China,which is a good material for understanding molecular mechanisms behind heavy metal pollutant tolerance in mangrove plants.In this study,based on the result of suppressive subtractive hybridization using rapid amplification of cDNA ends(RACE)technology,AmNramp3 gene was cloned using gene specific primers,with the full-length cDNA of 1 958 bp.We analyzed the expression of the AmNramp3 gene in leaf,stem and root after Cd treatment based on the quantative real-time PCR(qRT-PCR),and found the expression level in leaf was higher than in stem and root,while exhibited rapid response in stem and root.Additionally,the open reading frame(ORF)of the AmNramp3 gene was predicted and analyzed using bioinformatics,and results show that it is located in 1 bp to 1 527 bp and encodes 508 amino acids.Then the structure of the AmNramp3 protein was predicted and analyzed,and results show that the protein has no signal peptide,indicating that it is not a secretory protein.However,the AmNramp3 protein has a complex transmembrane structure,indicating that it is a transmembrane protein.Therefore,our analysis predicts that AmNramp3 encodes a transmembrane protein and the expression is rapidly changed under Cd stress,which is possibly related to the transmembrane transport function of heavy metal cadmium.This is helpful for us to understand the molecular mechanism of the AmNramp3 gene function in terms of heavy metal tolerance.