(College of the Environment & Ecology,Xiamen University,Xiamen 361102,China)
mangroves; water logging; high salinity; tolerance mechanism
DOI: 10.6043/j.issn.0438-0479.201612003
备注
红树林生境具有高盐、水淹低氧等特征,红树植物对高盐、水淹生境的适应性决定了其可以成功定植并占据相应的潮间带.红树植物可以通过各种机制耐受、抵抗或避免高盐、水淹,在海岸潮间带下生存.在生理及分子水平综述了红树植物适应水淹和高盐生境的研究进展,揭示了其独特的调控模式,为进一步研究其适应性机制提供了基础.
The unique characteristics of mangrove habitats are water logging,low oxygen and high salinity.To adapt to these intertidal environments,mangrove plants develop a set of adaptive mechanisms.The research progresses in physiological and molecular adaptation to water flooding and high saline habitats of mangrove plants are reviewed in this paper.Some unique tolerance regulations in mangrove plants are highlighted,which could provide a new direction for further studying adaptive mechanisms of mangrove plants to water logging and high saline habitats.
引言
红树植物是指能正常生长于热带、亚热带滨海潮间带或河上边缘的木本植物群落[1].作为存在于陆地和海洋潮间带的独特生物群落,红树植物可以促淤造陆抵消水面上升,从而降低因全球气候变暖而引起海平面升高对人类生存造成的威胁[2],其对于环境生态保护和社会经济发展的作用得到了广泛的认可.红树林生境复杂,自然状态下高盐、高温、强光、水淹对红树植物及其幼苗生长均具有显著影响.国内外学者对红树植物耐水淹和高盐适应性进行的大量研究表明:长期进化过程中,红树植物进化出一套适应于潮间带生长的耐水淹和高盐适应性机制[3-4].本文中将着重从生理和分子水平对近年来红树植物耐水淹和高盐适应性的研究进展进行论述.
1 红树植物的耐水淹适应性
2 红树植物的高盐适应性
3 结 语
作为生长在海岸潮间带的木本植物,红树植物的生境复杂,其最主要的特点是高盐和水淹缺氧.虽然不同种属的红树植物耐水淹和高盐的能力不尽相同,但是均能通过一系列的代谢途径来抵御环境中ROS对植物体内蛋白、膜脂类和核酸等细胞成分的伤害.目前在生理水平上对水淹和高盐胁迫下红树植物的光合作用特性、抗氧化酶活性、生物量变化有较多的研究,而对能直接与ROS反应,也可作为酶的底物出现在ROS清除机制中的物质如α-生育酚、类胡萝卜素和类黄酮等的研究,以及对于在可控温室下研究红树植物苗期生长、生物量积累与分配和植株Na
随着科学技术的发展,近年来关于红树植物水淹或高盐胁迫下的调控机制也开展了一些研究,从单个基因的转录表达分析到对白骨壤、秋茄、木榄等红树植物的转录组测序和蛋白质组测序,但是目前分子水平的研究还停留在胁迫诱导差异表达基因的筛选和相关基因的分析; 同时由于缺乏基因组信息,红树植物的转录组拼接和注释过程中可能存在较高概率的人为误差.对于红树植物仍有许多尚未解决和值得关注的重要问题,更全面地研究红树植物,深入了解其耐水淹和高盐适应性机理,对有效利用关键基因培育优良的作物品种具有重要的理论意义和应用价值.
2.1 红树植物的耐盐性红树植物生境的主要特征之一是高盐,其来源是浅海海水,潮水每天周期性浸润潮间带土壤,使土壤盐度增高,为NaCl型.在印度地区一年中夏季海水盐度最高,Kathiresan等[36]测得红树植物生长地——印度东南沿海皮恰瓦兰的河口潮间带土壤盐度冬季在1.0%~3.4%(质量分数,下同),夏季在1.12%~3.15%.红树植物不仅能适应盐度的动态变化,而且也能长期生长在高盐环境,形成一系列特定的形态结构来适应盐生环境[37].
根据红树植物有无盐腺,可将其分为泌盐红树植物和非泌盐红树植物[1].泌盐红树植物的叶片或茎表皮细胞可分化成盐腺以排除体内多余的盐分,维持体内一定浓度的盐分,从而减轻盐分对植物造成的伤害[1]; 非泌盐红树植物根细胞质膜的主要组成成分是三萜醇,三萜醇是一种植物次级代谢产物,会随着盐浓度的增加而增加,使盐离子不易通过细胞质膜,减少通过细胞膜的盐离子,降低木质部中盐分的含量[38].在根系的形态结构上,泌盐红树植物和非泌盐红树植物也明显不同:自然生长条件下,泌盐植物白骨壤根系疏水屏障内皮层只有单层细胞,随着外源盐浓度上升,从距离根尖20 mm处开始,内皮层细胞增加,出现双层细胞; 而非泌盐植物柱果木榄(B. cylindrica)在自然生长条件下,从距离根尖10 mm处开始就可以看到丰富的双层内皮层细胞[39].
红树植物作为专性盐生植物,在整个生命周期中需要一定浓度的盐,虽然在淡水或低盐下能存活生长,但在高盐下植物的生长发育会受到阻碍[40].秋茄幼苗在低盐(0%~2.0%)下,叶片数目增加,叶片含水量上升,叶片表面积增大,在高盐(2.0%~5.0%)下叶片发育受到抑制,气孔密度也低于低盐下的; 色素和单宁含量则呈现低盐抑制、高盐促进的规律[41].
2.2 红树植物的耐盐机制2.2.1 渗透调节作用较高的盐分在土壤中沉积,土壤中会产生一个低水势区域,致使植物根系难以从土壤中获得水分和营养,因此高盐会造成植物的生理性干旱,使之处于水分亏损的状态[42].植物在对盐渍的适应过程中,细胞会积累一定数量的可溶性有机物质,作为渗透调节物质进行渗透调节,以适应外界的低水势[43].
高盐下脯氨酸、甜菜碱和多醇等多种重要渗透调节物质在红树植物体内积累.对昆士兰地区23种红树植物的比较研究发现,松醇和甘露醇作为红树植物细胞中重要的相容性物质,提高了植物体内的渗透压,在盐分进入叶片或非原生质体时保持膨压,维持了细胞质中的水分[44]; 用核磁共振光谱鉴别得杯萼海桑(Sonneratia alba)中的相容性物质是甘露醇[45]; 小花木榄(B. parviflora)在400 mmol/L NaCl处理45 d后脯氨酸发生积累[46]; 600 mmol/L NaCl胁迫处理可使白骨壤叶片中甜菜碱、脯氨酸含量增加[47].其他重要的相容性物质,如一些含氮化合物(NCC)和小分子质量的碳水化合物,在高盐下也有变化.高盐下小花木榄叶片中淀粉含量减少,还原与非还原糖类增加,总氨基酸含量升高[46]; 桐花树在高盐下总糖含量降低,淀粉含量升高[47]; 高盐胁迫下,海莲(B. sexangula)叶片中的可溶性糖类含量随盐浓度升高而降低[48].
一些红树植物在高盐胁迫下,次级代谢物的含量也会发生变化.小花木榄在高盐处理45 d后积累多酚[46]; 在培养液中添加500 mmol/L NaCl,3 d后秋茄叶片中酚类、黄酮、花青素、原花青素和木质素含量升高[49]; 在高盐胁迫下,桐花树叶片中谷胱甘肽和多酚含量降低,但高盐胁迫的同时施加一定量的外源一氧化氮可以促进叶片中GSH和多酚等物质含量的升高[50].
2.2.2 离子转运与均衡在生理水平上,红树植物可以通过超滤等手段积累或隔离盐离子,从而限制细胞内盐含量[51].非泌盐红树植物拒盐效率是泌盐红树植物的10倍左右,泌盐红树植物(如白骨壤)主要通过根系内皮层中发达的凯氏带起作用,非泌盐红树植物(如秋茄和木榄)则是根细胞通过胚轴保存盐分或根系超滤作用拒绝盐离子进入植物体内[39,52]; 而角果木(Ceriops tagal(perr.)C. B. Rob)则通过根内皮层疏水屏障的增厚将90%~95%的盐分排除在体外[53].用同位素示踪法研究泌盐植物美洲红树在高盐下的离子吸收,标记48 h后,90%以上的36Cl-在根中,79%的22Na+在茎和胚轴中,叶片中只有0.2%的36Cl-且几乎没有22Na+[54].海莲悬浮细胞在低盐时积累Na+,高盐时排出Na+[48].X射线衍射研究显示,高盐胁迫下白骨壤叶片盐腺泌盐的增加与Na+和Cl-在根皮层中优先析出有关[55].红树植物Na+的排出可能是由细胞质膜上Na+/H+反向转运子(SOS1)完成的,SOS1利用细胞质膜上腺嘌呤核苷三磷酸(ATP)酶产生驱动力转运Na+,在一定盐度范围内,白骨壤与秋茄叶片和根中的ATP酶活性随着盐度的增大而增加,但过高的盐度则抑制ATP酶活性[56]; 向高盐胁迫处理下的白骨壤和秋茄施加一定浓度的外源一氧化氮,可以增加H+-ATP酶和Na+/H+转运子活性和相应基因表达量,促进白骨壤和秋茄在高盐下主动排盐,缓解盐离子对植物体的毒害[57-58].
2.2.3 抗氧化保护酶促和非酶促清除ROS途径在红树植物耐盐性中也起着重要作用[59].木榄和小花木榄在高盐处理下SOD活性急剧上升,当把木榄幼苗置于500 mmol/L NaCl中9 d,与对照相比SOD活性上升8.1倍[40]; 在白骨壤叶片中,SOD在高盐处理6周后活性升高[60].过氧化氢酶(CAT)参与植物体内过氧化氢的清除,在木榄属植物中,CAT活性随盐度的变化存在种间差异,如:小花木榄中CAT活性随着盐浓度增加而降低[61]; 木榄中CAT活性在250 mmol/L NaCl胁迫下降低,而在500 mmol/L NaCl胁迫下升高[40].在另一项研究中,高盐处理会引起白骨壤根和茎中POD活性升高[60].在Dasgupta等[62]的研究中,盐度的增加会引起木果楝(Xylocarpus granatum)、海漆(Excoecaria agallocha)、木榄、层孔银叶木(Heritiera fomes)、湄公河木果楝(X. mekonginsis)、大刺葵(Phoenix paludosa)和阿吉木(Aegialitis rotundifolia)的CAT和SOD活性增加.秋茄在高盐下启动抗氧化防御系统,通过提高SOD和POD活性来抵御高盐引起的氧化应激[63].除了抗氧化酶外,非酶类抗氧化剂在高盐胁迫下也存在相应的变化,如高盐胁迫下,小花木榄叶片抗坏血酸和谷胱甘肽含量水平下降[61].
2.3 红树植物耐盐性相关基因的表达及调控红树植物耐盐性与耐盐相关基因的表达和调控密切相关.表1列举了已报道在基因水平上对红树植物进行耐盐性研究的部分成果.通过建立高盐下秋茄cDNA文库及微阵列分析获得大量表达的基因,这些基因涉及光合作用、ROS清除和ROS代谢及平衡相关过程; 高盐胁迫状态下,与降低细胞内渗透势密切相关的水通道蛋白编码基因MIP、与蛋白质修复和降解错误折叠蛋白类相关的基因、与信号转导途径以及响应外界非生物刺激类相关的基因均上调表达,另外与离子转运和平衡相关的基因也高效表达[63].
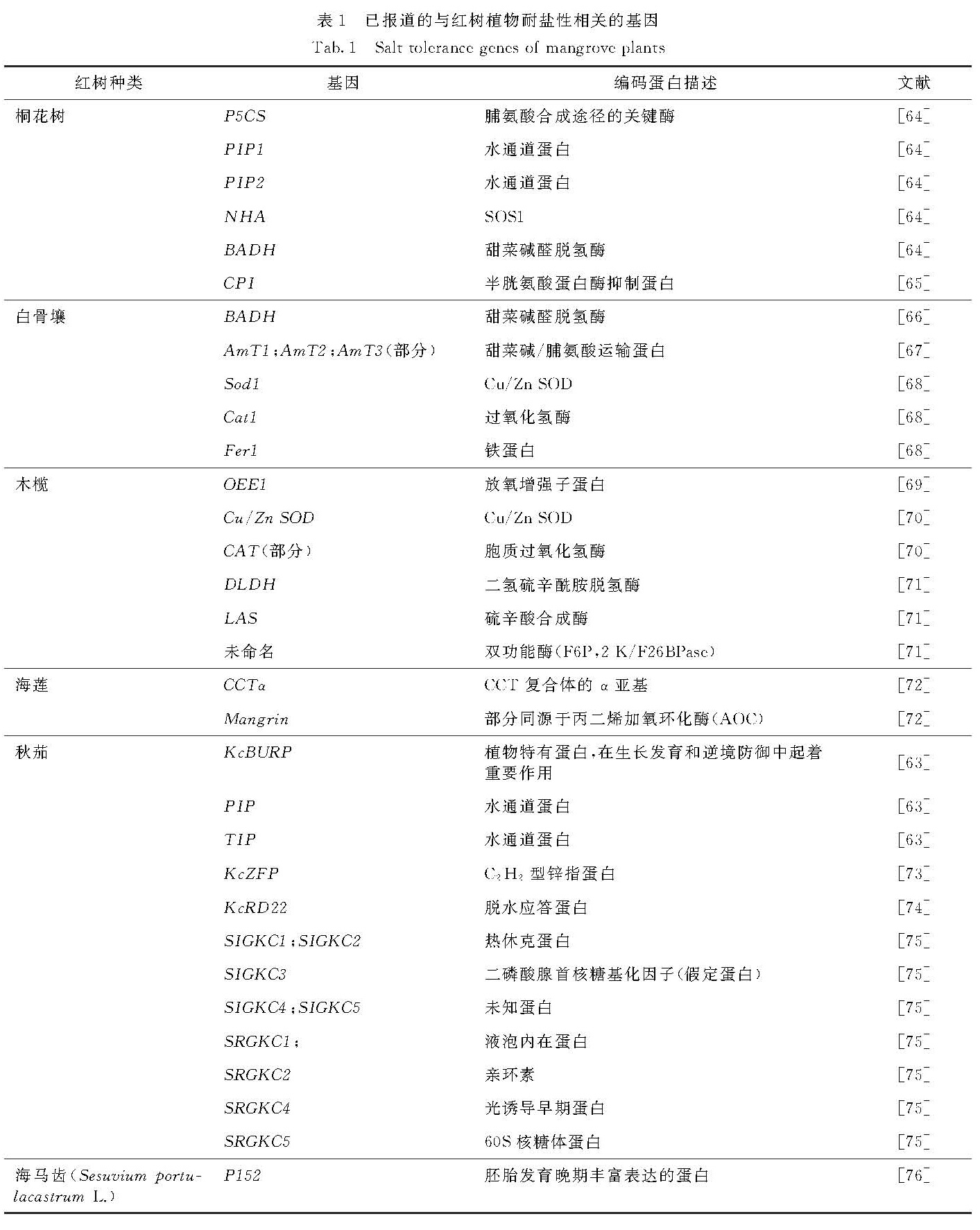
表1 已报道的与红树植物耐盐性相关的基因
Tab.1 Salt tolerance genes of mangrove plants以大肠杆菌(Escherichia coli)为宿主筛选小花老鼠簕(A. ebracteatus)的耐盐基因,通过组装拼接序列分析共筛选出107个与耐盐相关的候选基因,涉及参与应激反应功能蛋白的编码基因,与甘露醇脱氢酶、质体醛缩酶、分泌型POD、抗坏血酸POD和液泡H+-ATP酶活性变化相关的基因,以及与渗透平衡、离子转运和清除活性氧毒害密切相关的基因[77].
OEE1(oxygen evolving enhancer 1)蛋白含量上升有益于保持光系统Ⅱ(PSⅡ)的稳定和完整,提高光合效率.在500 mmol/L NaCl处理下,木榄PSⅡ外周OEE1蛋白表达增强[69].Takemura等[70]从木榄中克隆得到长度为153个氨基酸,编码液泡Cu/Zn-SOD的全长基因和编码CAT的部分基因序列,Northern杂交分析表明,液泡Cu/Zn-SOD基因的转录水平在NaCl处理后1~5 d内增加,但CAT基因表达水平的变化不明显.Miyama等[78]用500 mmol/L NaCl胁迫处理木榄后,微阵列分析获得865个差异表达基因,基因表达水平在盐处理6 h后发生变化,24 h后恢复以适应盐胁迫.Banzai等[71]用500 mmol/L NaCl处理木榄0,6,72和672 h后进行转录组测序,获得4 000个差异基因; 挑选6-磷酸-2-蛋白激/果糖-2(2-K/F26BPase)、磷酸酶2(F6P)、二氢硫辛酰胺脱氢酶(DLDH)和硫锌酸合成酶(LAS)的编码基因进行基因表达分析,在高盐处理6 h后2-K/F26BPase和F6P基因的表达快速上调,DLDH和LAS基因则在处理24 h后开始上调表达.
将白骨壤中参与甜菜碱合成的BADH基因转入大肠肝菌,转基因株系盐耐受水平增强[66].在烟草(Nicotiana glauca)中过表达秋茄C2H2型锌指蛋白编码基因KcZFP,过表达植株的耐盐性明显提高,脯氨酸水平远高于野生型,净光合作用速率受高盐胁迫的影响小于野生型植株,光系统在一定程度上得到了保护[73].同样,向烟草中转入秋茄KcRD22基因,解除高盐胁迫后,转基因烟草光合作用的恢复情况较野生型更好,可见KcRD22的过表达提高了烟草的耐盐性[74].
- [1] LIN P.Mangrove ecosystem in China[M].Beijing:Science Press,1999.
- [2] CALDERON H,WEEDA R,UHLENBROOK S.Hydrological and geomorphological controls on the water balance components of a mangrove forest during the dry season in the Pacific coast of Nicaragua[J].Wetlands,2014,34(4):685-697.
- [3] SCHOLANDER P F,DAM L V,SCHOLANDER S I.Gas exchange in the roots of mangroves[J].American Journal of Botany,1995,42(1):92-98.
- [4] 茹巧美,郑海雷,肖强.红树植物耐盐机理研究进展[J].云南植物研究,2006,28(1):78-84.
- [5] 张乔民,张叶春.华南红树林海岸生物地貌过程研究[J].第四纪研究,1997,4:344-353.
- [6] 黄敏参.福建漳江口滨海湿地植物水分利用格局及其生态学意义[D]:厦门:厦门大学,2012:1-114.
- [7] 利容千.植物逆境细胞及生理学[M].武汉:武汉大学出版社,2002:1-467.
- [8] 廖宝文,郑德璋,郑松发,等.红树植物秋茄造林技术的研究[J].林业科学研究,1996,9(6):586-592.
- [9] 宋鹏华,曾其伟,商敬哲,等.植物对水淹胁迫响应的研究进展[J].蚕业科学,2013,39(1):160-165.
- [10] 游惠明.秋茄幼苗对盐度、淹水环境的生长适应[J].应用生态学报,2015,26(3):675-680.
- [11] MCKEE K L,CAHOON D R,FELLER I C.Caribbean mangroves adjust to rising sea level through biotic controls on change in soil elevation[J].Global Ecology and Biogeography,2007,16(5):545-556.
- [12] HE B Y,LAI T,FAN H,et al.Comparison of flooding-tolerance in four mangrove species in a diurnal tidal zone in the Beibu Gulf[J].Estuarine Coastal & Shelf Science,2007,74(1):254-262.
- [13] CASTANEDA-MOYA E,TWILLEY R R,RIVERA-MONROY V H,et al.Patterns of root dynamics in mangrove forests along environmental gradients in the Florida coastal Everglades,USA[J].Ecosystems,2011,14(7):1178-1195.
- [14] ELLISON A M,FARNSWORTH E J.Seedling survivorship,growth,and response to disturbance in Belizean Mangal[J].American Journal of Botany,1993,80(10):1137-1145.
- [15] 林玲玲,黄丽,仇建标,等.长期水淹胁迫下的北移秋茄形态结构和生理变化[J].科技通报,2016,32(5):56-60.
- [16] 伍卡兰,彭逸生,郑康振,等.五种红树植物通气组织对人工非潮汐生境的响应[J].生态学报,2010,30(24):6927-6934.
- [17] LU W Z,CHEN L Z,Wang W Q,et al.Effects of sea level rise on mangrove Avicennia population growth,colonization and establishment:evidence from a field survey and greenhouse manipulation experiment[J].Acta Oecologica,2013,49:83-91.
- [18] 陈鹭真,林鹏,王文卿.红树植物淹水胁迫响应研究进展[J].生态学报,2006,26(2):586-593.
- [19] PEZESHKI S R,DELAUNE R D,MEEDER J F.Carbon assimilation and biomass partitioning in Avicennia germinans and Rhizophora mangle seedlings in response to soil redox conditions[J].Environmental & Experimental Botany,1997,37(2):161-171.
- [20] 廖宝文,邱凤英,管伟,等.尖瓣海莲幼苗对模拟潮汐淹浸时间的适应性研究[J].林业科学研究,2009,22(1):42-47.
- [21] KRAUSS K W,MCKEE K L,LOVELOCK C E,et al.How mangrove forests adjust to rising sea level[J].New Phytologist,2014,202(1):19-34.
- [22] KATHIRESAN K,BINGHAM B L.Biology of mangroves and mangrove ecosystems[J].Advances in Marine Biology,2001,40(1):81-251.
- [23] LANE D,MICHALOWSKI W,STEPHENSON R,et al.Integrated systems analysis for marine site evaluations and multicriteria decision support for coastal aquaculture[J].Aquaculture Innovation & Social Transformation,2008,17:255-264.
- [24] DAS S K,PATRA J K,THATOI H.Antioxidative res-ponse to abiotic and biotic stresses in mangrove plants:a review[J].International Review of Hydrobiology,2016,101(1/2):3-19.
- [25] DREW M C.Oxygen deficiency and root metabolism:injury and acclimation under hypoxia and anoxia[J].Plant Biology,1997,48:223-250.
- [26] STROMMER J.The plant ADH gene family[J].Plant Journal for Cell & Molecular Biology,2011,66(1):128-142.
- [27] CHEN L Z,WANG W Q,LIN P.Photosynthetic and physiological responses of Kandelia candel L. Druce seedlings to duration of tidal immersion in artificial seawater[J].Environmental and Experimental Botany,2005,54(3):256-266.
- [28] 薛鑫,张芊,吴金霞.植物体内活性氧的研究及其在植物抗逆方面的应用[J].生物技术通报,2013(10):6-11.
- [29] YE Y,TAM N F Y,WONG Y S,et al.Growth and physiological responses of two mangrove species(Bruguiera gymnorrhiza and Kandelia candel)to waterlogging[J].Environmental and Experimental Botany,2003,49(3):209-221.
- [30] 何斌源,赖廷和,王文卿,等.梯度淹水胁迫下全日潮海区秋茄幼苗的生长和生理反应[J].海洋通报,2007,26(2):42-49.
- [31] YOUSSEF T,SAENGER P.Anatomical adaptive strategies to flooding and rhizosphere oxidation in mangrove seedlings[J].Australian Journal of Botany,1996,44(3):421-426.
- [32] YANG Y,YANG S,LI J,et al.Transcriptome analysis of the Holly mangrove Acanthus ilicifolius and its terrestrial relative,Acanthus leucostachyus,provides insights into adaptation to intertidal zones[J].BMC Genomics,2015,16(1):1-12.
- [33] 郑琼,马旭俊,杨传平.硫氧还蛋白(Trx)的研究进展[J].分子植物育种,2006,4(S2):78-82.
- [34] 张留恩,廖宝文,管伟.模拟潮汐淹浸对红树植物老鼠簕种子萌发及幼苗生长的影响[J].生态学杂志,2011,30(10):2165-2172.
- [35] 李赞.红树植物秋茄幼苗叶片应对水淹胁迫的生理响应及差异蛋白质组学分析[D]:厦门:厦门大学,2016:1-73.
- [36] KATHIRESAN K.A review of studies on Pichavaram mangrove,southeast India[J].Hydrobiologia,2000,430(1):185-205.
- [37] REEF R,LOVELOCK C E.Regulation of water balance in mangroves[J].Annals of Botany,2015,115(3):385-395.
- [38] OKU H,BABA S,KOGA H,et al.Lipid composition of mangrove and its relevance to salt tolerance[J].Journal of Plant Research,2003,116(1):37-45.
- [39] KRISHNAMURTHY P,JYOTHI-PRAKASH P A,QIN L,et al.Role of root hydrophobic barriers in salt exclusion of a mangrove plant Avicennia officinalis[J].Plant Cell & Environment,2014,37(7):1656-1671.
- [40] WANG W,YAN Z,YOU S,et al.Mangroves:obligate or facultative halophytes? A review[J].Trees,2011,25(6):953-963.
- [41] 李元跃,黎中宝,潘文,等.盐度对秋茄幼苗的生长和某些生理生态特性的影响[C]∥福建省科协第八届学术年会.莆田:福建省科学技术协会,2008:137-141.
- [42] 杨启良,张富仓,刘小刚,等.植物水分传输过程中的调控机制研究进展[J].生态学报,2011,31(15):4427-4436.
- [43] 靖姣姣,张颖,白志英,等.盐胁迫对小麦代换系渗透调节物质的影响及染色体效应[J].植物遗传资源学报,2015,16(4):743-750.
- [44] POPP M,LARHER F,WEIGEL P.Osmotic adaptation in Australian mangroves[J].Vegetatio,1985,61(1):247-253.
- [45] YASUMOTO E,ADACHI K,KATO M,et al.Uptake of inorganic ions and compatible solutes in cultured mangrove cells during salt stress[J].In Vitro Cellular & Developmental Biology-Plant,1999,35(1):82-85.
- [46] MOHANTY P.Defense potentials to NaCl in a mangrove,Bruguiera parviflora:differential changes of isoforms of some antioxidative enzymes[J].Journal of Plant Physiology,2004,161(5):531-542.
- [47] ZHU Z,ZHANG R F,LIU T W,et al.Solute accumulation and osmotic adjustment characteristics of the mangrove Avicennia marina under NaCl-induced salinity stress[J].Botanica Marina,2011,54(3):335-341.
- [48] 郑文教,林鹏.盐度对红树植物海莲幼苗的生长和某些生理生态特性的影响[J].应用生态学报,1992,3(1):9-14.
- [49] WANG L,PAN D,LÜ X,et al.A multilevel investigation to discover why Kandelia candel thrives in high salinity[J].Plant Cell & Environment,2016,39(11):2486-2497.
- [50] CHEN J,XIAO Q,WANG C,et al.Nitric oxide alleviates oxidative stress caused by salt in leaves of a mangrove species,Aegiceras corniculatum[J].Aquatic Botany,2014,117(1):41-47.
- [51] ZHENG W J,WANG W Q,LIN P.Dynamics of element contents during the development of hypocotyls and leaves of certain mangrove species[J].Journal of Experimental Marine Biology,1999,233:248-257.
- [52] LAWTON J R,TODD A,NAIDOO D K.The structure of the roots of the mangroves,Avicennia marina and Bruguiera gymnorrhiza,in relation to ion uptake[J].New Phytologist,1981,88(4):713-722.
- [53] AZIZ I,KHAN M A.Experimental assessment of salinity tolerance of Ceriops tagal seedlings and saplings from the Indus delta,Pakistan[J].Aquatic Botany,2001,70(3):259-268.
- [54] MALLERY C H,TEAS H J.The mineral ion relations of mangroves Ⅰ.Root cell compartments in a salt excluder and a salt secreter species at low salinities[J].Plant & Cell Physiology,1984,25(7):1123-1131.
- [55] PARIDA A K,DAS A B,MITTRA B.Effects of salt on growth,ion accumulation,photosynthesis and leaf anatomy of the mangrove,Bruguiera parviflora[J].Trees,2004,18(2):167-174.
- [56] ZHAO Z Q,ZHENG H L,ZHU Y G.Changes of plasma membrane ATPase activity,membrane potential and transmembrane proton gradient in Kandelia candel and Avicennia marina seedlings with various salinities[J].Journal of Environmental Sciences,2004,16(5):742-745.
- [57] CHEN J,XIAO Q,WU F,et al.Nitric oxide enhances salt secretion and Na+ sequestration in a mangrove plant,Avicennia marina,through increasing the expression of H+-ATPase and Na+/H+ antiporter under high salinity[J].Tree Physiology,2010,30(12):1570-1585.
- [58] CHEN J,XIONG D Y,WANG W H,et al.Nitric oxide mediates root K+/Na+ balance in a mangrove plant,Kandelia obovata,by enhancing the expression of AKT1-Type K+ channel and Na+/H+ antiporter under high salinity[J].PLoS One,2013,8(8):1-11.
- [59] JITHESH M N,PRASHANTH S R,SIVAPRAKASH K R,et al.Antioxidative response mechanisms in halophytes:their role in stress defence[J].Journal of Genetics,2006,85(3):237-254.
- [60] CHERIAN S,REDDY M P,PANDYA J B.Studies on salt tolerance in Avicennia marina(Forstk.)Vierh.:effect of NaCl salinity on growth,ion accumulation and enzyme activity[J].Indian Journal of Plant Physiology,1999(4):266-270.
- [61] PARIDA A K,DAS A B,MOHANTY P.Defense potentials to NaCl in a mangrove,Bruguiera parviflora:differential changes of isoforms of some antioxidative enzymes[J].Journal of Plant Physiology,2004,161(5):531-542.
- [62] DASGUPTA N,NANDY P,SENGUPTA C,et al.Protein and enzymes regulations towards salt tolerance of some Indian mangroves in relation to adaptation[J].Trees,2012,26(2):377-391.
- [63] 侯佩臣.高盐胁迫下红树植物秋茄的表达谱微阵列分析及差异表达基因的功能解析[D]:北京:北京林业大学,2010:1-129.
- [64] FU X,HUANG Y,DENG S,et al.Construction of a SSH library of Aegiceras corniculatum under salt stress and expression analysis of four transcripts[J].Plant Science,2005,169(1):147-154.
- [65] 傅新晖.桐花树耐盐基因的克隆、功能表达及其适应性进化的研究[D].广州:中山大学,2006:1-149.
- [66] HIBINO T,MENG Y L,KAWAMITSU Y,et al.Molecular cloning and functional characterization of two kinds of betaine-aldehyde dehydrogenase in betaine-accumulating mangrove Avicennia marina(Forsk.)Vierh.[J].Plant Molecular Biology,2001,45(3):353-363.
- [67] WADITEE R,HIBINO T,TANAKA Y,et al.Functional characterization of betaine/proline transpor-ters in betaine accumulating mangrove[J].Biological Chemistry,2002(277):18373-18382.
- [68] JITHESH M N,PRASHANTH S R,SIVAPRAKASH K R,et al.Monitoring expression profiles of antioxidant genes to salinity,iron,oxidative,light and hyperosmotic stress in the high salt toletant grey mangrove,Avicennia marina(Forsk.)Vierh.by mRNA analysis[J].Plant Cell Reporter,2006(25):865-876.
- [69] SUGIHARA K,HANAGATA N,DUBINSKY Z,et al.Molecular characterization of cDNA encoding oxygen evolving enhancer protein 1 increased by salt treatment in the mangrove Bruguiera gymnorrhiza[J].Plant and Cell Physiology,2000,41(11):1279-1285.
- [70] TAKEMURA T,HANAGATA N,DUBINSKY Z,et al.Molecular characterization and response to salt stress of mRNAs encoding cytosolic Cu/Zn superoxide dismutase and catalase from Bruguiera gymnorrhiza[J].Trees,2002,16(2):94-99.
- [71] BANZAI T,HERSHKOVITS G,KATCOFF D J,et al.Identification and characterization of mRNA transcripts differentially expressed in response to high salinity by means of differential display in the mangrove,Bruguiera gymnorhiza[J].Plant Science,2002,162(4):499-505.
- [72] YAMADA A,SAITOH T,MIMURA T,et al.Expression of mangrove allene oxide cyclase enhances salt tolerance in Escherichia coli,yeast,and tobacco cells[J].Plant Cell Physiology,2002(43):903-910.
- [73] 于瑞,胡月,黄旭新,等.过表达秋茄C2H2型锌指蛋白基因KcZFP提高烟草耐盐性[J].基因组学与应用生物学,2013,32(2):149-158.
- [74] 黄旭新,侯佩臣,丁明全,等.秋茄KcRD22基因的克隆与功能分析[J].基因组学与应用生物学,2011,30(4):273-280.
- [75] HUANG W,FANG X D,LI G Y,et al.Cloning and expression analysis of salt responsive gene from Kandelia candel[J].Biologia Plantarum,2003,47(4):501-507.
- [76] ZENG H C,DENG L H,ZHANG C F.Cloning of salt tolerance-related cDNAs from the mangrove plant Sesuvium portulacastrum L.[J].Journal of Integrative Plant Biology,2006,48(8):952-957.
- [77] NGUYEN P D,HO C L,HARIKRISHNA J A,et al.Functional screening for salinity tolerant genes from Acanthus ebracteatus Vahl using Escherichia coli as a host[J].Trees,2007,21(5):515-520.
- [78] MIYAMA M,HANAGATA N.Microarray analysis of 7029 gene expression patterns in burma mangrove under high-salinity stress[J].Plant Science,2007,172(5):948-957.
1.1 红树林生长带与潮汐水位的关系红树植物的分布和生长受潮汐水淹的影响,根据生物学和生态学特性,红树植物在潮间带的分布呈现出低滩带、中潮带和高潮带的带状分布特征,使红树植物具有向海性生态序列现象[5].红树植物水分利用保守,潮汐活动可明显改变红树林生态系统的水热平衡及植物水分利用:潮汐浸淹使秋茄(Kandelia obo-vata)和白骨壤(Avicennia marina)树干茎流密度以及日蒸腾量下降,红树林生态系统的初级生态系统生产力、净生态系统交换和生态系统也分别降低了6.56%、6.84%和12.08%; 同时为了抵御潮汐活动带来的水淹和高盐胁迫,减少体内水分消耗,红树植物水分利用保守[6].另外潮汐也影响红树植物胚轴着床和生长,持续水淹时间过长,植物呼吸和光合作用受到抑制,植物体内生理过程发生改变,土壤产生厌氧反应,一些化学物质被还原成有毒化合物被红树植物吸收,从而导致林木生长弱小,对环境胁迫抵抗力差,最终逐渐死亡[7].廖宝文等[8]在海南东寨港、广东湛江和深圳福田三地沿海潮间带营造秋茄试验林的实验结果显示:造林6个月后,在东寨港和湛江的成活率均达到90%以上,在福田的成活率为74.1%; 且在福田浅水带(低于平均海面3 cm左右)3年后成活率和保存率分别为76.9%和62.2%,效果最佳,在深水带(低于平均海面51 cm左右)3年后成活率和保存率分别为47.8%和3.2%,效果最差.该结果说明潮淹深度对秋茄幼苗成活率和保存率影响极为明显.
1.2 水淹对红树植物的影响1.2.1 水淹对红树植物形态及组织结构的影响对大多数中生植物而言,水淹会限制其有氧呼吸,抑制其能量的产生,一些植物在水淹胁迫下还会出现根系分蘖增加、不定根增生和形成根系气腔的现象[9],影响其生长.而对于生长在潮间滩涂的红树植物来说,能适应一定深度和时长的潮汐水淹[10],但水淹过深且时间过长时其形态结构也会发生变化,如出现叶片变小、生长受阻和根系变短等现象[11].He等[12]在搭建于广西英罗湾红树林外裸滩的试验平台上设置了8个海面高程组,研究了全日潮海区潮汐水淹对桐花树(Aegiceras corniculatum)、白骨壤、木榄(Bruguiera gymnorrhiza)和红海榄(Rhizophora stylosa)幼苗的生长和生理影响,结果表明:小高程(低于平均海面20~40 cm)和大高程(高于平均海面20~30 cm)均促进木榄和红海榄幼苗茎高度的增长,中等高程(低于或高于平均海面10 cm)则起抑制作用; 中等高程组幼苗茎节数稍少,叶数、叶面积与叶保存率均随水淹程度加大而增加,所有高程组均有一些幼苗死亡,高程越小幼苗存活率越低,由73.6%下降到35.0%; 而对于桐花树和白骨壤,大高程抑制幼苗茎高度的增长,叶数、叶面积与叶保存率均随水淹程度加大而急剧减小,不同高程下幼苗均存活.Castaneda-Moya等[13]在大沼泽地国家公园设置多个固定样点进行观测采样,发现由于水淹频率的增加使黑皮红树(A. germinans)、拉关木(Laguncularia racemosa)和美洲红树(R. mangle)3种红树植物生长受到抑制,细根生物量下降.Ellison等[14]对美洲红树和白骨壤幼苗在不同高度潮滩的生长进行了调查,发现生长在平均潮位的幼苗一年后存活率分别为56%和47%,最低潮位和最高潮位生长的幼苗一年后均死亡; 高潮位上的美洲红树和白骨壤幼苗萌发初期生长较快,当植株成苗后生长速率下降,地下部分的比例升高; 与之相反,低潮位生长的植株矮小,分枝和叶片少; 而在平均潮位的植株,各生长指标均比低潮位或高潮位生长的植株高10%~20%,相对生长率高3%~23%.
对秋茄的水淹处理试验也显示其组织结构和显微结构出现变化,表现为:叶片角质层增厚,海绵组织细胞间隙变大,叶肉细胞体积变小,叶绿体淀粉粒堆积,原生质体出现明显的质壁分离; 叶绿体结构出现轻微膨胀,基粒片层与片层间较稀疏,部分叶绿体膜遭到破坏; 根皮层形成大量气生根,上皮细胞壁角质化,皮层中形成大量通气组织[15].潮汐生境下华南地区5种红树植物优势种根中通气组织大小比较结果是红海榄>秋茄>白骨壤>木榄>桐花树; 茎和叶中通气组织大小比较则显示白骨壤最大,桐花树最小[16].
1.2.2 水淹对红树植物生理生化的影响1)对光合作用的影响
在水淹环境下,植物根系缺氧会影响叶片的光合特性[9].伍卡兰等[16]的研究发现,白骨壤等5种红树植物在水淹初期气孔关闭,随着水淹时间延长,与光合作用相关的酶活性降低,叶绿素含量下降.Lu等[17]对一年生白骨壤幼苗进行不同水淹时长和淹没深度的室内模拟实验,发现随着水淹时长的增加,白骨壤幼苗叶片的光合作用速率、光合作用电子传递和水分利用的效率均降低,总生物量下降; 全株水淹和仅根系水淹相比,全株水淹对白骨壤幼苗负面影响更大,幼苗全株水淹12 h后出现死亡.Ellison等[14]发现,低潮位的美洲红树幼苗与中潮位的相比,气孔密度增加7%,但气孔导度降低11%,光合作用速率减小6%.其他相关研究也指出,水淹状态下白骨壤、拉关木、美洲红树、秋茄和木榄等红树植物叶片气孔关闭,气孔导度降低,光合作用速率减小,总生物量降低[10,18].但也有个别红树植物水淹状态下的叶绿素a与b比值不降反升,如木榄叶片的叶绿素含量会随着水淹高度的上升而增加,长时间的水淹使叶绿素a与b比值上升[19]; 尖瓣海莲(B. sexangula var. rhynchopetala)叶片的叶绿素a与b比值也会随着水淹时间的延长而上升[20].
2)对呼吸作用的影响
在水淹环境下,红树植物体内的O2、CO2等气体扩散受到限制,乙烯积累增加,水下叶片光截获能力下降,光合作用减弱,植物细胞内线粒体活性氧(ROS)产生,从而引发氧化胁迫[21].受潮汐影响,红树植物根系经常处于被淹没状态,缺氧明显[22],从而通过一系列自适应结构和代谢途径来抵抗缺氧[23],表现为形态特化,生长出大量气生根,暴露在空气中进行气体交换,同时,体内产生的次级代谢物能有效清除有氧或无氧呼吸过程中产生的过量CO2[24].
水淹胁迫下,湿生植物优先选择乙醇发酵途径[7],在丙酮酸脱氢酶复合体(PDC)和乙醇脱氢酶(ADH)作用下,产生乙醇这类更易扩散到胞外的物质[25-26].Przeshki等[19]发现随着水淹程度的加剧,白骨壤和美洲红树幼苗根系中的ADH活性大幅度增加.Chen 等[27]也发现,随着水淹时间的增加,秋茄幼苗ADH活性增加,在水淹处理4~6 h时,根系的ADH活性达到最高值.
3)ROS积累及抗氧化系统活性变化
在水淹环境下,植物体内ROS大量增加,蛋白质、膜脂、DNA及其他细胞组分严重受损,而植物体内的抗氧化酶类能减轻ROS积累造成的危害,增加植物抗逆性[28].水淹胁迫下,木榄叶片超氧化物歧化酶(SOD)无明显变化,过氧化物酶(POD)活性变化明显[29].何斌源等[30]研究发现:秋茄幼苗根系SOD和POD活性随着海面高程的增加而升高,在360 cm海面高程(高于平均海面1 cm)时达到最大值,而后开始降低; 叶片中SOD和POD活性随着海面高程的增加而降低.Youssef等[31]发现,耐水淹能力强的红树植物白骨壤和美洲红树幼苗根际氧化酶含量高于木榄和桐花树,同时耐水淹能力强的红树植物根表皮细胞的孔隙度也更大,对根际氧气有更强的束缚力,以此缓解水淹胁迫引起的细胞缺氧,减轻ROS积累.
1.3 红树植物适应水淹生境的分子调控红树植物的自然生境受周期性海水浸淹的影响,研究其适应水淹生境的调控机理对于保护红树植物的繁衍生长具有现实意义.
Yang等[32]对红树植物老鼠簕(Acanthus ilicifolius)及其近缘陆生种刺苞老鼠簕(A. leucostachyus)进行了比较转录组分析,发现其中有12个基因与DNA和细胞膜损伤修复以及ROS清除过程相关,基因Ail_c48553_g1_i1和Ail_c56385_g1_i1参与植物中重要的抗氧化剂还原型谷胱甘肽(GSH)的合成,另有一个基因Ail_c51326_g2_i1的同源基因编码硫氧还蛋白CDSP32,在逆境胁迫下参与植物防御胁迫引起的膜脂过氧化[33].张留恩等[34]发现:短期的潮汐淹没对老鼠簕种子萌发没有影响,但当淹没时间大于每天15 h时种子萌发率降低; 挖掘转录组数据后发现了7个与植物种子萌发和胚胎发育相关的基因,其中Ail_CL5647Contig1基因的同源基因编码拟南芥(Arabidopsis thaliana)泛素连接酶蛋白BRIZ2,参与形成不同的泛素连接酶复合体,这些复合体是种子萌发和萌发后生长所必需的; 其他3个基因Ail_CL3016Contig1、Ail_CL3870Contig1和Ail_c54714_g1_i1在种子胚胎发育中发挥重要的作用,大大提高了老鼠簕在水淹环境下繁殖的适应能力.
李赞[35]运用双向电泳技术研究了水淹4 d处理下秋茄叶片蛋白质组的变化:经过基质辅助激光解吸飞行时间质谱鉴定,分析得到差异蛋白质46个,其中大多数定位于叶绿体内,少数分布在线粒体、高尔基体、细胞核、细胞液、过氧化物酶体中,主要与光合作用和能量产生、物质代谢过程、氧化还原平衡、激素信号响应相关; 同时发现与脱落酸(ABA)信号转导途径密切相关的硫氧还蛋白CDSP32和转录因子myc2 bHLH表达量均下调.由此推测,秋茄可能通过ABA信号转导途径调控植株采取“逃离策略”以提高其耐水淹能力.
