红树林是泛热带的海岸木本植物群落,天然分布在我国东南沿海的海南、广东、广西、福建等省份以及台湾、香港和澳门地区; 浙江省在1957年成功引种秋茄(Kandelia obovata)[1].红树植物都是嗜热种类,低温是限制红树林向两极扩展的主要因子[6].我国处于世界红树林天然分布区的北缘,不同地域自然分布的红树林植物群落的种类多样性随纬度增高而递减[7].依据最低月均温的等温线图和红树植物天然分布的北缘,林鹏和傅勤[8]把我国红树植物的耐寒性划分为7个等级,由Ⅰ~Ⅶ级耐寒能力逐渐减弱(表1).
表1 中国红树植物耐寒性等级序列
Tab.1 Cold tolerant classes of mangrove species in China
注:引自文献[8],经修改.
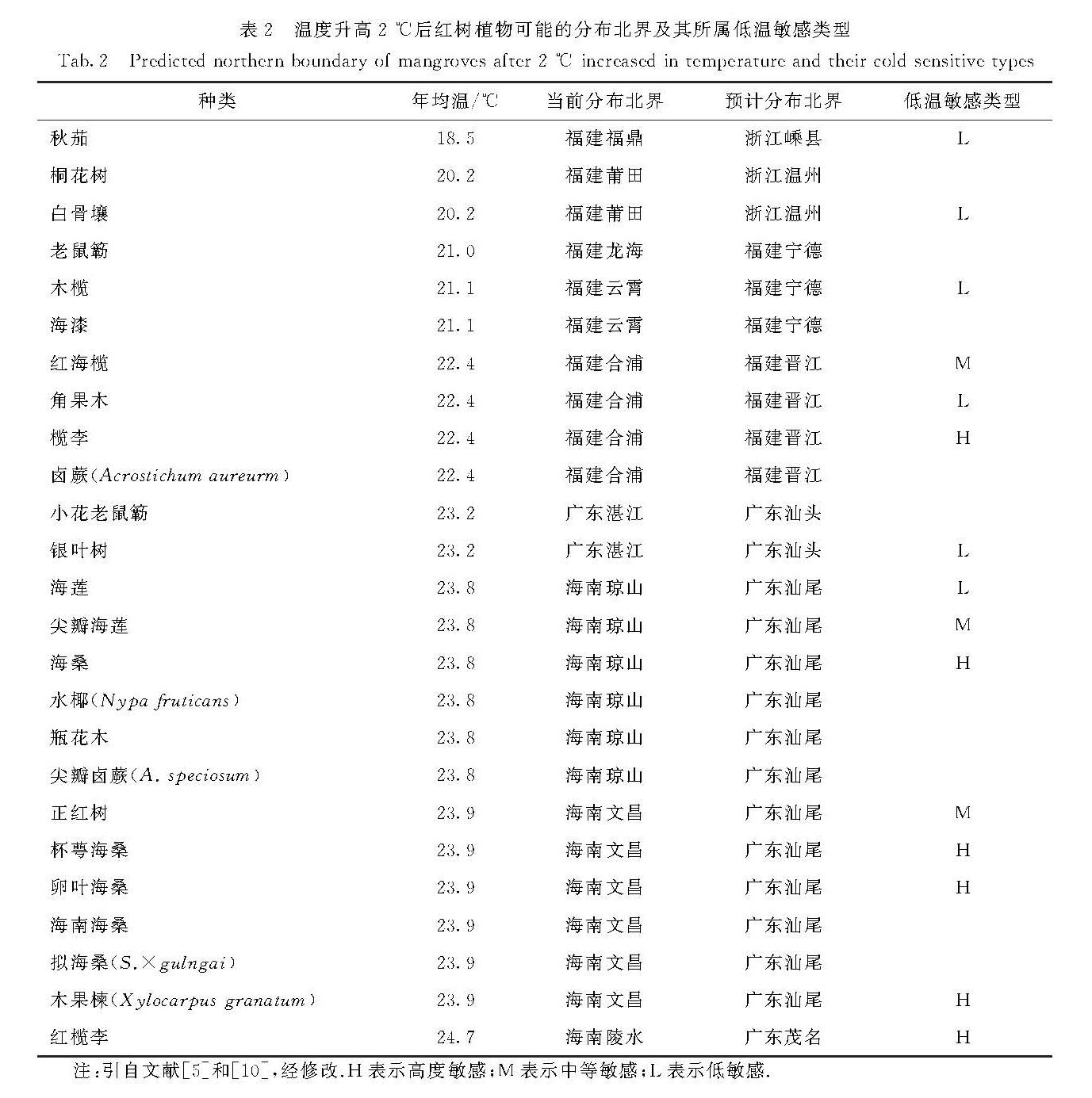
根据这一耐寒性等级序列,我国最耐寒的红树植物是秋茄,其自然分布区的纬度最高(福建福鼎),生境的最低月均温为8.4 ℃,也是最早向北引种到浙江的种类; 而红榄李和水芫花的分布仅限于海南岛南端.张娆挺和林鹏[9]从植物区系的角度出发,把红树植物分为抗低温广布种、嗜热广布种和嗜热窄布种,将广布性与耐寒性紧密结合.然而,全球气温升高将给红树植物的两极扩展提供机会.基于我国红树林分布现状和IPCC对全球气温升高的预测,陈小勇和林鹏[5]对未来气温升高2 ℃后的红树林地理分布范围进行了理论预测:各种红树植物的分布区可能在纬度上平均北扩2.5°; 我国红树林分布的北界将从福建福鼎扩展至浙江嵊县附近,引种的北界达杭州湾; 升温后,浙江省可能有3个红树种类(表2).
表2 温度升高2 ℃后红树植物可能的分布北界及其所属低温敏感类型
Tab.2 Predicted northern boundary of mangroves after 2 ℃ increased in temperature and their cold sensitive types
注:引自文献[5]和[10],经修改.H表示高度敏感; M表示中等敏感; L表示低敏感.
在全球范围内,气候变暖的过程也将伴随着极端低温发生频率的增加,同时由于不同红树种类的耐寒能力不同,低温将对北移的种类产生不同程度的损害.围绕低温的胁迫,林鹏等开展了一系列红树林植物耐寒性的生理生态学研究[10-12]:自1980年开始,卢昌义、杨盛昌等从海南采集木榄、红海榄、海莲、尖瓣海莲、角果木、正红树的成熟胚轴,在室外盆栽至高为10~20 cm后再移植至福建九龙江红树林引种园; 之后,沈瑞池把一些较为耐寒的种类(如白骨壤和桐花树)从九龙江口移植到莆田、宁德、福鼎、温州等地.通过对比发现:桐花树可以引种到福鼎和宁德,且其繁殖体能在宁德萌发和生长; 白骨壤北移后较难成活; 秋茄能在温州生长,但无法自然更新; 木榄在九龙江口能自然萌发、生长并繁育后代; 红海榄、海莲、尖瓣海莲虽能在九龙江口生长,但冬季的低温使其成活率低于40%; 角果木和正红树在引种后经历第一个冬季时即全部死亡.卢昌义等[12]对海南岛引种到九龙江口的木榄、海莲和尖瓣海莲在低温敏感阶段(2年内幼树期)的生物量、光合水分利用等进行跟踪研究,发现它们对于高纬度的低温有不同程度的生理生态适应能力.林鹏等[13]通过盆栽试验,对秋茄、桐花树、白骨壤、海莲、木榄和尖瓣海莲6种红树植物的临界低温进行了生理生态学指标测定,其结果与野外引种试验的结果一致,即气温10 ℃是这些红树植物的生物学临界温度.
杨盛昌[10]通过采集海南琼山东寨港的主要红树植物种类,利用电导率法测定其耐寒能力,发现海南东寨港主要红树种类冬季半致死温度介于-2.3~-6.8 ℃之间,因此认为:红树科植物耐寒能力较强,多数种类属于低温敏感性相对较弱的L类型; 而海桑科和楝科红树种类耐寒能力较弱,属于低温敏感性较强的H类型(表2).杨盛昌和林鹏[14]进一步比较了不同纬度红树林区、同一红树群落不同滩面高程以及同一植株树冠不同位置的耐寒能力; 从生理生态水平揭示了红树植物的耐寒适应表现,并提出红树植物叶片耐寒能力的种群分化、季节变化和部位差异是其趋异适应的结果,有益于其生存和繁育.
这一系列研究工作在一定程度上为当时红树林全球变化响应研究提供了数据积累和参考依据.此后十多年的几次大寒潮,从野外实践上再次证明了这一红树林耐寒性等级序列的适用性.例如,2008年初,我国南方19个省经历了50年一遇的持续低温雨雪冰冻天气,极端气候对华南沿海各省的红树林区造成了不同程度的危害.陈鹭真等[15]在2008年寒潮过后的1个月内,对我国南方各省红树林区的10个代表性地点进行红树植物伤害程度的系统调查发现:极端低温对红树林的危害极为显著,特别是在低纬度的海南省、广西省和广东湛江等地; 但在纬度较高的福建省,本地红树种类秋茄、桐花树和白骨壤及引种的木榄,由于长期适应于冬季较低的气温或在种植前经过耐寒锻炼,具有较强的耐寒能力.这一结果基本符合杨盛昌和林鹏[14]的红树植物耐寒性等级序列.此次调查还发现当极端低温正值夜间退潮时,红树林受寒害更为显著[15].冬季海水温度20 ℃被认为是红树林分布的临界水温[16],因此,对红树植物低温胁迫的研究也从气温进一步延伸到气温和水温的相互作用.Wang等[17]探讨了福建福鼎(秋茄天然分布北界)的秋茄群落在冬季霜冻持续下叶片衰老和养分循环的机制,提出了海水对于霜冻的保温作用.陈鹭真等[18]通过室内控制模拟冬季低温和夜间低潮对无瓣海桑幼苗的影响,进一步验证了极端低温发生时夜间高潮对幼苗起到了较好的保温作用,缓解了低温对幼苗生长和叶片的生理伤害.目前,国内学者对红树植物耐寒性机制的探讨集中在光合作用和叶绿素荧光、养分利用、低温保护机理等方面[14,18-20].
未来的全球变暖趋势将有利于红树植物生长[4-5].邓燕瑜[21]和胡娜胥[22]通过控制实验研究发现:秋茄、无瓣海桑(S. apetala)等红树植物对未来气温升高的适应方式有所不同.胡娜胥[22]依据陈小勇和林鹏[5]的推测,对气温升高2 ℃后秋茄和无瓣海桑的地理分布进行推测,并获得与之相似的结果.史小芳[23]通过野外调查也发现:作为我国最耐寒的红树植物,秋茄在低纬度区域(海南省)的分布面积小,整体长势差,植株高度低,其生长在一定程度上受到夏季高温的抑制; 这与夏季高温抑制秋茄的最大电子传递速率、破坏叶绿体结构,进而抑制光合作用密切相关.因此,在未来气温升高和极端低温的协同作用下,红树植物的生理生态学响应和红树林分布范围的两极化值得进一步关注.